





Abstract
Anthropogenic disturbances of wildlife, such as noise, human presence, hunting activity, and motor vehicles, are becoming an increasing concern in conservation biology. Fireworks are an important part of celebrations worldwide, and although humans often find fireworks spectacular, fireworks are probably perceived quite differently by wild animals. Behavioral responses to fireworks are difficult to study at night, and little is known about the negative effects fireworks may have on wildlife. Every year, thousands of tons of fireworks are lit by civilians on New Year’s Eve in the Netherlands. Using an operational weather radar, we quantified the reaction of birds to fireworks in 3 consecutive years. Thousands of birds took flight shortly after midnight, with high aerial movements lasting at least 45 min and peak densities measured at 500 m altitude. The highest densities were observed over grasslands and wetlands, including nature conservation sites, where thousands of waterfowl rest and feed. The Netherlands is the most important winter staging area for several species of waterfowl in Europe. We estimate that hundreds of thousands of birds in the Netherlands take flight due to fireworks. The spatial and temporal extent of disturbance is substantial, and potential consequences are discussed. Weather radar provides a unique opportunity to study the reaction of birds to fireworks, which has otherwise remained elusive.
INTRODUCTION
The impact of anthropogenic disturbance on wildlife is a major concern in conservation biology (e.g., Bowles 1995; Gill and Sutherland 2000; Buckley 2004). Facultative responses such as changes in behavior due to diverse anthropogenic disturbances including human presence, transportation (e.g., low-flying aircraft, motor boats), hunting, and noise have been studied in terrestrial as well as marine animals (e.g., Bélanger and Bédard 1989; Andersen et al. 1996; Constantine et al. 2004; St Clair et al. 2010). Many of these of disturbances can be considered unpredictable labile, or transient, perturbation factors that cause a facultative response at the individual level (e.g., Rykiel 1985; Wingfield 2003). Numerous studies have focused on the behavioral and physiological responses as well as potential longer term consequences of such disturbances to long-lived animals like water birds during different periods in their annual routine (e.g., Burger 1981; Bélanger and Bédard 1990; Madsen 1995; Béchet et al. 2004; Klaassen et al. 2006). For example, noise due to low-flying aircraft caused flocks of staging snow geese to stop feeding and take flight, reducing the amount of time spent foraging, and potentially resulting in a significant energetic cost (Bélanger and Bédard 1989, 1990).
For centuries, firework displays have been an important part of celebrations worldwide. Although generally perceived by humans as spectacular, the unexpected loud noises and bright lights fireworks produce are probably a source of disturbance for many species of domestic and wild animals. For example, studies have shown that fireworks are a major traumatic event for dogs (Landsberg et al. 2003; Levine et al. 2007). Similarly, pyrotechnics and gas exploders are used to actively disperse wildlife and are applied worldwide as an effective measure in bird strike prevention (Gilsdorf et al. 2002; Cook et al. 2008). Yet, the effects fireworks may have on wildlife are hardly known.
In many countries around the world, fireworks are lit on New Year’s Eve and national holidays; these may be huge centralized displays as well as more dispersed fireworks lit by private individuals. Numerous studies worldwide have documented the negative impact fireworks have on public health through pollution, physical injuries, auditory, and visual trauma (e.g., Plontke et al. 2002; Drewnick et al. 2006; Moreno et al. 2007; Saadat et al. 2010). In the Netherlands, civilians are allowed to light their own fireworks on New Year’s Eve and do so on a massive scale in nearly all inhabited areas. It is estimated that 10.8 million kg of fireworks are ignited each year in the Netherlands (van der Maas et al. 2010), resulting in numerous accidents. The Netherlands is a densely populated country with areas set aside for nature conservation occurring in close proximity to inhabited areas. We propose that firework displays are an anthropogenic disturbance, which can be considered an indirect labile perturbation factor (a rapid event that does not affect the animal directly; following Wingfield 2003) for various animals. Due to the proximity between wildlife and people in the Netherlands, we expect fireworks to have an observable effect on wildlife. However, as fireworks are mainly lit at night, the effect on animals and its extent is difficult to study in general.
In this study, we used an operational weather radar to quantify the spatial and temporal extent of disturbances caused by large-scale recreational fireworks in the Netherlands on New Year’s Eve. Radar has the advantage that it is capable of detecting birds at large distances and independent of light conditions (Bruderer 1997; Gauthreaux and Belser 2003). Although radar has been used extensively to study bird, bat, and insect movements (e.g., Gauthreaux and Belser 2003; Horn and Kunz 2008; Chapman et al. 2011), to the best of our knowledge, this is the first time that radar was used to quantify the immediate response of animals to human disturbance.
METHODS
Study area
As illustrated in Figure 1a, the radar is located close to several important fresh water lakes, grasslands, and river floodplains, which are used by thousands of wintering water birds (Hustings et al. 2009). We focused our quantitative analysis on the area above the wetlands and shallow lakes included in the Natura 2000 nature conservation site Oostelijke Vechtplassen (lat 52.2°N, long 5.05°E, Anonymous 2010), located 10–18 km from the radar. Monthly winter counts in this area show that mainly wintering Greylag (Anser anser L., 6000–7000 individuals) and Greater white-fronted geese (Anser albifrons S., 2500–3000 individuals) sleep on the lake (ca. 560 birds/km2), and Wigeon (Anas penelope L., ca. 11 000 individuals) and mallards (Anas platyrhynchos L.,) roost on the lake during the day and feed in the surroundings at night (Hustings et al. 2009).
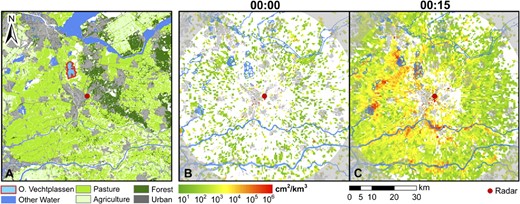
Study area and radar reflectivity measured on New Year’s Eve, 2008; (a) Overview of the study area in the Netherlands, including general land cover. The radar at De Bilt (lat 52.103°N, long 5.179°E) is indicated by a red dot and the blue lines and polygons show major rivers, lakes, and wetlands in the area. The lake and wetland area Oostelijke Vechtplassen (lat 52.2°N, long 5.05°E) is indicated by a red polygon. (b) Overlaid radar reflectivity (square centimeter/cubic kilometer) at 1 January 2009 00:00 LT(GMT +1). The area where the beam center exceeds 750 m altitude is shaded in gray, as birds mainly fly up to about 500 m (Figure 2e), not many echoes are expected outside this area. (c) Radar reflectivity (square centimeter/cubic kilometer) at 1 January 2009 00:15 LT. Red and orange areas indicate hot spots of bird activity predominantly over water bodies. Supplementary Material, Supplementary movie 1 shows the reflectivity in 5-min intervals for region shown in this figure.
Radar measurements
According to Dutch law, consumer fireworks can be sold between 29 and 31 December. Fireworks can be lit for recreational use from 31 December 10:00 to 1 January 2:00 and are typically concentrated in the first 30 min after midnight. We used an operational C-band weather radar (SELEX-SI, Neuss-Rosellen, Germany) in De Bilt, the Netherlands (lat 52.103°N, long 5.179°E, 44 m above mean sea level) to monitor echoes by flying birds at 5-min intervals, in various altitude bands within a radius of 25 km from the radar. We used data from 30 December 00:00 to 3 January 00:00 LT (Local Time, GMT +1), in the years 2007–2010 (i.e., 3 years). The method for quantifying bird movement was extensively validated on this radar in previous studies (Holleman et al. 2008; van Gasteren et al. 2008; Dokter et al. 2011). Bird quantification methods were identical to those described previously (Dokter et al. 2011), with 2 modifications needed for this study. First, the bird reflectivity estimate was based only on resolution volumes directly above the Oostelijke Vechtplassen (water body area 16 km2). Focusing our analysis on the Oostelijk Vechtplassen had the advantage that it is within the standard measurement window of the algorithm, where the radar beam is sufficiently narrow to probe the altitude profile of flying birds. Furthermore, although radar may also register reflectivity caused by fireworks in the lowest air layers (e.g., up to 100 m), we strongly limited the amount of scattering contaminations by fireworks at the lowest altitudes by confining our analysis to water bodies in a protected reserve. The second modification entailed increasing the reflectivity factor threshold, which classifies high reflectivity as precipitation clutter, from 20 to 35 dBZ to include the full range of reflectivity values due to birds during peak disturbance. We obtained a vertical profile of bird reflectivity corresponding to the mean beam height of each elevation scan at the study site (0.1, 0.2, 0.3, 0.5, 0.8, 1.1, 1.6, 2.1, 2.8, 3.4, 4.4, 6.2, and 8.3 km above ground level, respectively).
Estimating bird densities
Reflectivity (square centimeter/cubic kilometer) and Vertically Integrated Reflectivity (VIR, square centimeter/square kilometer) can be converted into volumetric and height-integrated bird densities respectively, by dividing by the radar cross-section (RCS, square centimeter) for the typical bird at the site (Dokter et al. 2011). The seasonally averaged RCS during passerine migration equals 11 cm2 (Dokter et al. 2011). Based on annual winter bird counts in the radar area (Hustings et al. 2009), we assumed that the majority of birds are large waterfowl during this period of time. Using this information, we obtained an order of magnitude estimate for the RCS of waterfowl by scaling the passerine RCS to the relatively larger body area for ducks and geese. Assuming a relative body volume of 1:10:50 for passerines:ducks:geese and assuming the body area-to-volume fraction scales as a sphere, we obtained a rough estimate for the RCS of 50 cm2 for ducks and 150 cm2 for geese, which is similar to values described in other studies (Bruderer and Joss 1969; Bruderer 1997).
RESULTS
We detected very little bird movement right before midnight on New Year’s Eve, as shown by the low radar reflectivity in Figure 1b. In contrast, only several minutes after 00:00 LT, we detected massive bird movement when fireworks were lit in inhabited areas all over the country; this can be seen by the very high radar reflectivity in the full surveyed area of the radar (Figure 1c). We observed hot spots of activity over lakes, wetlands, and river floodplains (red areas in Figure 1c, Supplementary Material, Supplementary Movie 1), many of which are designated as Natura 2000 sites (Notenboom et al. 2006; Anonymous 2010). These sites hold large numbers of water birds which rest and forage in the area (Hustings et al. 2009). By examining the spatial reflectivity patterns in 5-min intervals, we observed that birds took off and dispersed from lake areas and wetlands (Supplementary Material, Supplementary Movie 1) and in some cases clearly moved away from densely populated areas (Supplementary Material, Supplementary Movies 1 and 2; note that the wind direction was from the north for this year and movement of the reflectivity peak is clearly toward the north, indicating that this is not due to reflectivity from fireworks). Measurements were impeded directly above urban areas, as well as within 10 km distance from the radar, because ground clutter was severe in these areas and no valid reflectivity measurements passed the radar signal processor’s clutter filtering.
We calculated the radar reflectivity at 5-min intervals for the air volume above the wetlands and shallow lakes included in the Natura 2000 site Oostelijke Vechtplassen (lat 52.2°N, long 5.05°E, Figure 1a) and show the time series for each year in Figure 2a–d. We observed short peaks in reflectivity on the days and nights before and after New Year’s Eve; however, these did not exceed a VIR of 4000 cm2/km2 (Figure 2a–c). We observed a very abrupt and strong increase in density each year beginning on 1 January at 00:05 LT and reaching a maximum between 00:15 and 00:25 LT (Figure 2d). Densities then decreased and returned to base values by 01:30 LT. Peak density was highest on 1 January 2009, and the maximum VIR was 1.4 × 104 cm2/km2. Birds flew up to approximately 500 m altitude (Figure 2e). Flight altitudes increased rapidly during the first 15–20 min after midnight, and then slowly decreased, with the main disturbance period lasting about 45 min. The same temporal and altitude pattern was seen over other lakes and wetlands in the radar area.
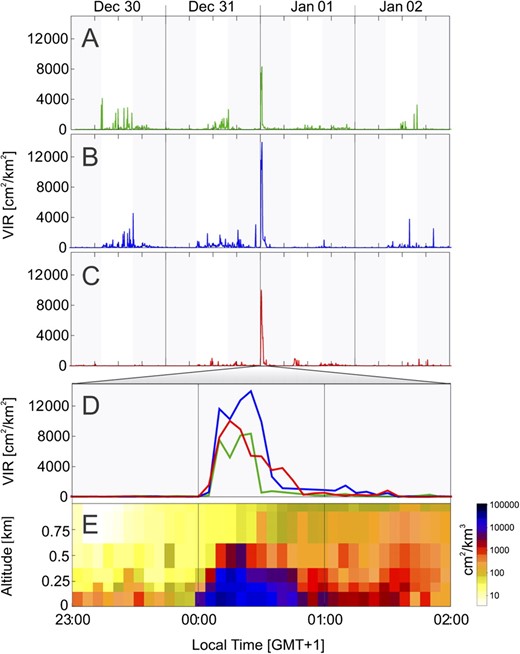
Time series of bird movements over Loosdrechtse Plassen. VIR (square centimeter/square kilometer) from 30 December 00:00 to 3 January 00:00 for (a) 2007/2008, (b) 2008/2009, and (c) 2009/2010. (d) VIR (square centimeter/square kilometer) from 31 December 23:00 to 1 January 02:00 (2007/2008 green, 2008/2009 blue, and 2009/2010 red). (e) Altitude density profile from 31 December 2008 23:00 to 1 January 2009 02:00 over Oostelijke Vechtplassen. Altitude (kilometer) is shown on the y axis and time on the x axis. Colors represent measured reflectivity (square centimeter/cubic kilometer). Gray shaded areas in 2a–d indicate the time between sunset and sunrise.
DISCUSSION
Numerous studies have shown that animals change their behavior and show physiological responses to loud abrupt noises (e.g., Weisenberger et al. 1996; Delaney et al. 1999, see also Bowles 1995 for review). In this study, fireworks were a source of significant disturbance to birds, causing them to take flight directly after midnight flying up to altitudes of several hundred meters. The activity of birds closely follows the use of fireworks. In 2007 and 2008 (blue and green lines, Figure 2d), a first peak in activity can be seen just after midnight with a second peak 15 min later. Considering the temporal and spatial patterns simultaneously, we attribute this to a first group of birds taking off, with a proportion of the birds dispersing out of the measurement area and then a second group of birds taking off, potentially due to a second burst of fireworks. Similarly, a study in Germany on atmospheric particle concentrations due to fireworks on New Year’s Eve showed closely matching temporal patterns of particle concentrations attributed to a first bout of fireworks, a pause due to the exchange of good wishes called “the Champagne dip,” followed by a more prolonged period of fireworks (Drewnick et al. 2006). Directly around cities, close to where most fireworks were lit, radar reflectivity remained low compared with protected reserves, where no fireworks were lit. The contribution of fireworks measured reflectivity is therefore relatively small, and a contribution from fireworks particles advected from urban areas into the protected reserves can be safely neglected. The low spikes observed during the day, just before New Year’s Eve (Figure 2a–c) could probably be attributed to smaller scale disturbance of birds when fireworks were sporadically lit. In 2007, for example, thousands of geese were visually observed at count stations flying in numerous directions (Waanders and Troost 2010).
We estimate that 100 birds/km2 are in the air over this area at peak disturbance (assuming a RCS of 100 cm2). Using the same conversion, the red/orange hot-spot areas in Figure 1c correspond to 1000 birds/km2. The Netherlands is the most important winter staging area for several species of waterfowl in Europe, with approximately 2 million geese observed during peak winter counts (Hustings et al. 2009). If we extrapolate this disturbance outside of the radar measurement area, we expect that hundreds of thousands of waterfowl that regularly overwinter in the Netherlands are disturbed and take flight as an immediate behavioral response to New Year’s Eve fireworks.
Birds flew up to altitudes of several hundred meters, in contrast to daily local movements, which normally occur below 100 m (Dirksen et al. 1994; Gauthreaux and Belser 2003; Shamoun-Baranes et al. 2006). The spatiotemporal patterns indicate that individual birds flew several kilometers before settling again and may even remain in the air for more than 30 min. The immediate escape response requires birds to temporarily stop their “nonessential” activities, and birds may enter what is sometimes called an emergency life-history stage (e.g., Wingfield et al. 1998; Wingfield 2003). Several of the short-term consequences of taking flight would, in this study, include the energetic cost of flight (e.g., Pennycuick 2008), disruption of sleep (e.g., Amlaner and McFarland 1981; Lima et al. 2005), resting or foraging (e.g., Bélanger and Bédard 1989; Béchet et al. 2004), and a potential initial reduction in intake rates following resettlement, particularly if birds need to adjust to a new foraging site. Birds may also become disorientated during such evasive flights at night, particularly as visibility can deteriorate during extensive firework activity (Drewnick et al. 2006; Beijk et al. 2009). For example, intense smog was measured in several urban and even rural areas on New Year’s Eve 2008/2009 due to the combination of intense fireworks, mist, and low wind speeds (Drewnick et al. 2006; Beijk et al. 2009). Furthermore, disturbance may also have a negative effect on the immunocompetence of individuals (Tarlow and Blumstein 2007). Social cues such as flock size may also affect the magnitude of the response as shown in several studies about geese (e.g., Bélanger and Bédard 1989; Béchet et al 2004). As the majority of species measured in this study were slow-living birds, we expect a stronger emergency reaction to disturbance than might be observed for smaller fast living species, such as passerines (e.g., Ricklefs and Wikelski 2002; Weimerskirch et al. 2002; Bisson et al. 2009, 2011).
Although we do not expect fireworks to be directly lethal to birds, confounding factors, such as disorientation, or flying in inclement weather normally avoided could potentially result in mortality. A recent event which has attracted a great deal of media attention is the sudden and mass mortality of thousands of red-winged blackbirds that fell out of the sky in Beebe Arkansas on New Year’s Eve 2010. Similar to our study, Next Generation Weather Radar images (http://abcnews.go.com/Health/radar-images-show-startled-birds-taking-flight-arkansas/story? id=12781951) show that birds left their roosts between rain showers in several pulses on New Year’s Eve. A great deal of speculation surrounds this event, birds may have been scared into the air due to fireworks or other loud and abrupt noises, becoming disoriented and crashing into obstacles, suffering from stress, encountering inclement weather once in the air or a combination of effects and died. Without such a disturbance, the birds would probably have remained in their roost. Additional reports of birds mysteriously falling from the sky can be found in the media right around New Year’s Eve. Our study is just one example of a clear and large-scale behavioral response of birds to fireworks, a very specific form of disturbance. We believe that as awareness increases and more quantified observations are made, such events can be linked to a behavioral response of birds to loud and unpredictable noises and may not be so rare.
This study quantifies the temporal and spatial scale of immediate response of birds to fireworks; more work is clearly needed to assess the longer term consequences at the individual and population levels (e.g., Gill and Sutherland 2000; Wingfield 2003; Béchet et al. 2004; Wikelski and Cooke 2006), especially in and around areas designated for nature conservation, such as Natura 2000 sites (Anonymous 2000). The weather radar used in this study is part of an operational weather radar network (OPERA, Operational Programme for the Exchange of weather RAdar information) (Dokter et al. 2011) and as such can be a powerful tool for monitoring large movements (e.g., due to disturbance or migratory flights) across political boundaries and on a continental scale (Gauthreaux and Belser 2003; Shamoun-Baranes et al. 2008; Dokter et al. 2011). Although radar enables us to quantify the movements of birds in flight and at night, we can expect that other wildlife, not measured by the radar are also affected. Biotelemetry and field endocrinology can help further improve our understanding of individual behavioral and physiological response and the energetic cost of such disturbances in a broad range of animals (e.g., Cooke et al. 2004; Ropert-Coudert and Wilson 2005; Walker et al. 2005; Bisson et al. 2009), and an integration of techniques to study responses at different scales, at the individual and population level, could be even more promising (Robinson et al. 2010).
SUPPLEMENTARY MATERIAL
Supplementary material can be found at http://www.beheco.oxfordjournals.org/.
FUNDING
BiG Grid infrastructure for e-Science (www.biggrid.nl).
We thank S. Shinneman for assistance with the figures. We thank SOVON, The Dutch centre for field ornithology (www.sovon.nl) for providing winter bird count data. We thank F. Liechti and one anonymous reviewer for their helpful comments on an earlier version of this manuscript.

{kind=link}
{kind=link}