





Abstract
A new longevity record has been established for a free-living bat—41 years for a male Brandt's bat, Myotis brandtii. The exceptional longevity of bats generally, and this species in particular, should make bats of special interest for researchers studying mechanisms of slow aging.
A male Brandt's bat (Myotis brandtii) has recently been recaptured in the wild 41 years after it was originally banded, making it the world's longevity champion among small mammals (Figure 1). In terms of its longevity quotient, which is longevity standardized by body size (1), M. brandtii lives 9.8 times longer than expected—the highest value reported for any mammal, even those maintained in captivity. The fact that this animal could survive this long in the wild indicates very well-preserved function. That is, the animal had to have well-maintained high frequency hearing to detect prey with its echolocation, as well as fly fast enough, and maneuver with enough agility, to capture prey and avoid predators on a daily basis.
In a several-year period around 1962, 1544 individual bats were banded in several cave colonies of M. brandtii in the Biryusa karst region of Siberia in Russia. From this original marked population, at least 67 animals—all males—lived longer than 20 years (Table 1). Due to a scarcity of field data (there were no speleological expeditions in the region between 1990 and 2000), the numbers of animals living to between 28 and 38 years of age may be unrepresentative.
The general pattern for mammals is that larger species are longer-lived than smaller species (1). However, bats are a major exception to this pattern, with longevity of more than 20 years now documented for 22 bat species, more than 30 years for six species (2), and now more than 40 years for one species (Figure 2). Although several long-lived bat species are large (300–1000 grams), most are small. Thirteen species in the genus Myotis, ranging in size from 7 to 25 grams, have been documented to live at least 20 years in the wild. M. brandtii weighs only 6–6.7 grams before hibernation and 5–6.2 grams after hibernation. Khritankov and Ovodov (3) previously reported a maximum longevity record of 38 years for Brandt's bat. However, 3 years later the same bat was recaptured during hibernation.
A substantial period of yearly hibernation has been advanced as one cause of the exceptional longevity of bats. Indeed, M. brandtii hibernates from late September until the middle of June in Siberia. However, both hibernating and nonhibernating species of bats are long-lived. In a comparative analysis of bat longevity, the strongest correlate of longevity was reproductive rate, with short life being associated with more young bats produced per year. The next strongest correlate was hibernation, with hibernating bats living, on average, 6 years longer than nonhibernating bats. Latitude (associated with length of hibernation) and habit of cave roosting were also correlated with longevity (2).
It is not clear why all long-lived individuals are male, but the same pattern has been reported in another exceptionally long-lived species, the little brown bat (Myotis lucifugus, W. H. Davis, personal communication with S. N. A., 2002). Males spend longer in hibernation than do females, who have to awaken earlier to join maternity colonies. Also, because bat offspring are exceptionally large (about 25% of maternal body size), pregnant females are likely to be less agile in flight and may be more susceptible to predation. Female bats also face considerable energetic demands during lactation as they suckle their young until they are almost the size of adults (4). One or more of these factors could be involved in the apparent sex difference in longevity.
What other than sex might distinguish long-lived individuals from their shorter-lived colony mates? Very anecdotally, long-lived animals seem to choose hibernation sites within caves that are highly buffered from the outside environment, and they also seem exceptionally placid, not struggling exceptionally hard when being handled and returning to hibernation very quickly. This could potentially indicate that they are particularly deep or long hibernators. Longer hibernators have been shown to live longer than shorter hibernators in Turkish hamsters (5).
Of interest to gerontologists, of course, are the mechanisms that enable bats to survive so long and maintain their ability to function at a high level physically. One possibility is that they are particularly resistant to oxidative damage. M. lucifugus, which lives up to 34 years in the wild, has been reported to produce fewer reactive oxygen species per unit of oxygen consumed in heart, brain, and kidney compared with two similar-sized, but much shorter-lived, mammal species (short-tailed shrew, white-footed mouse). No differences between species were found in superoxide dismutase activity (6). Unfortunately, no measures of age-related oxidative damage to tissues have yet been reported for any species of bat.
We are currently investigating whether there is some feature of cultured fibroblasts that might be informative about the long lives of bats. Fibroblasts isolated from the skin of M. lucifugus experience less DNA damage when exposed to a range of γ-irradiation doses than do similar cells from mice (Figure 3). M. lucifugus fibroblasts also appear to repair DNA damage more quickly (A. J. P., S. N. A., unpublished data, 2005).
Although it is premature to claim at this point that either reduced oxidative damage or stress-resistant DNA is “why” M. lucifugus live up to 34 years in the wild, these findings are provocative. Little is known of basic biology of bat cells. Our results at least suggest that bats might have exceptional defense mechanisms against cellular damage.
M. brandtii might also be fascinating for understanding the interaction of genes and environment in aging, as other populations of the same species are apparently much shorter-lived. For instance, in the Leningrad region of Russia, M. brandtii is not apparently long-lived, possibly due to a different ecological history. Near St. Petersburg, Russia, M. brandtii is reported to live no more than 8.5 years (7). Moreover, during 50 years of research in the Baltic region, Masing and colleagues (8) reported that maximum life span of M. brandtii in the region never exceeded 18.5 years. Although winter conditions are slightly warmer around St. Petersburg (hibernation occurs only from September until May), there might also be higher predation pressure or more competition with other bat species. Siberia could be viewed as “safe island” compared with central Europe, where animals could live longer if the cold winter could be tolerated. Unfortunately, only old data for the longevity records of M. brandtii in the Leningrad region are available.
We hope that this report will stimulate interest among biogerontologists to consider bats seriously as study organisms in a comparative approach to aging research.
Decision Editor: James R. Smith, PhD
![Myotis brandtii roosting in a cave (A), and “facing” the camera (B). [Photos by Alexander Khritankov.]](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/biomedgerontology/60/11/10.1093/gerona/60.11.1366/2/m_grna-60-11-17-f01.gif?Expires=1716365253&Signature=pJ3WwujJeXi3ByV3CLRN8RBQtqic6GaF5yx6eyULyahrFhczikYLVdBg2cimOVwuYeUuc4QCQgH2s~UkdGrs8FofUBYxcxG~3qLJ5yLrF-csNp0sRNmJbctkGSmfIMcGTHjktUCteScQCVC3Kmuj2QTJVV1WveCxVKaXBdgmKULXAI4cj5fYezaFt-Mpq4cV5qUKz4~vMFySH-7gLX-~cCgsaQf3qTpRcx4M~R1FsiiaWm8jb0xnycydg9JZt98MoMT3u5imf7LseZzSIcHVhHuIc6tzLxk0zXAi6ji31CiTwAyzOpm31lC7eAN3b-lGisM54oqUlXz2x3cpd2gteg__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Myotis brandtii roosting in a cave (A), and “facing” the camera (B). [Photos by Alexander Khritankov.]
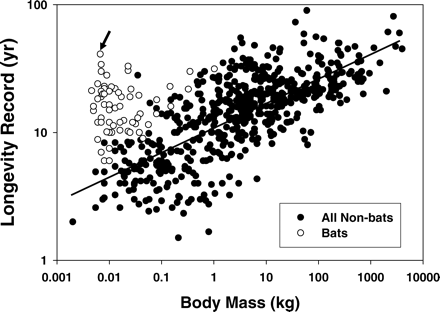
Longevity of bats compared with other mammal species. Note that some bat species do not seem excessively long-lived for their body size relative to other mammals. However, most longevity records for bats were accidentally acquired rather than systematically gathered. Therefore, record longevities in some species may be nonrepresentative because very small numbers of individuals have been marked and monitored. Arrow points to data point for Myotis brandtii
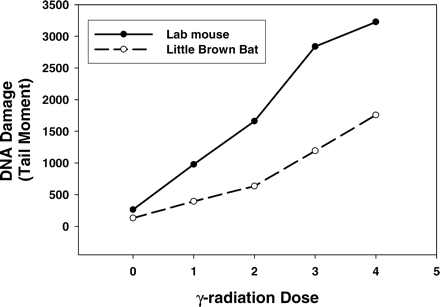
Greater resistance to DNA damage in the little brown bat, Myotis lucifugus, compared with a laboratory mouse. DNA damage was measured with alkaline single-cell gel electrophoresis technique–comet assay. This assay measures strand breaks and abasic sites. Tail moment is a measure of total DNA damage. Two hundred cells were analyzed for each data point. VisCOMET software (Impuls GmbH, Gilching, Germany) was used in analysis. Median values are presented because the distribution of individual cell values is not normally distributed. Cells were cultured in Dulbecco's modified Eagle medium with 10% fetal bovine serum, streptomycin, and penicillin, in 5% CO2 and 3% O2 at 37°C
Number of Survivors (All Males) to at Least the Age Indicated From 1544 Bats (of Both Sexes) Originally Banded.
| Age | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 33 | 38 | 39 | 41 |
| No. alive | 67 | 56 | 54 | 42 | 41 | 34 | 25 | 19 | 5 | 4 | 4 | 3 | 2 | 2 | 1 |
| Age | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 33 | 38 | 39 | 41 |
| No. alive | 67 | 56 | 54 | 42 | 41 | 34 | 25 | 19 | 5 | 4 | 4 | 3 | 2 | 2 | 1 |
Note: Modified from (3).
Number of Survivors (All Males) to at Least the Age Indicated From 1544 Bats (of Both Sexes) Originally Banded.
| Age | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 33 | 38 | 39 | 41 |
| No. alive | 67 | 56 | 54 | 42 | 41 | 34 | 25 | 19 | 5 | 4 | 4 | 3 | 2 | 2 | 1 |
| Age | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 33 | 38 | 39 | 41 |
| No. alive | 67 | 56 | 54 | 42 | 41 | 34 | 25 | 19 | 5 | 4 | 4 | 3 | 2 | 2 | 1 |
Note: Modified from (3).
This research was partially supported by U.S. Public Health Service grant AG-022873 from the National Institute on Aging.
References
Austad SN, Fischer KE. Mammalian aging, metabolism, and ecology: evidence from the bats and marsupials.
Khritankov AM, Ovodov ND. Longevity of Brandt's bats (Myotis brandtii Eversmann) in central Siberia.
Hayssen V, Kunz TH. Allometry of litter mass in bats: maternal size, wing morphology, and phylogeny.
Lyman CP, O'Brien RC, Greene GC, Papafrangos ED. Hibernation and longevity in the Turkish hamster Mesocricetus brandti.
Brunet-Rossinni AK. Reduced free-radical production and extreme longevity in the little brown bat (Myotis lucifugus) versus two nonflying mammals.
Strelkov PP. Ecological surveillance of bats during winter hibernation (Chiroptera, Vespertilionidae). Annals Institute of Zoology, USSR. 1971;21–30.


{kind=link}
{kind=link}
{kind=link}