





Abstract
Declines in monarch butterfly populations have prompted investigation into the sensitivity of their milkweed host plants to land-use change. Documented declines in milkweed abundance in croplands have spurred efforts to promote milkweeds in other habitats. Nevertheless, our current understanding of milkweed populations is poor. We used a long-term plant survey from Illinois to evaluate whether trends in milkweed abundance have caused monarch decline and to highlight the habitat-management practices that promote milkweeds. Milkweed abundance in natural areas has not declined precipitously, although when croplands are considered, changes in agricultural weed management have led to a 68% loss of milkweed available for monarchs across the region. Midsuccessional plant communities with few invasive species provide optimal milkweed habitat. The augmentation of natural areas and the management of existing grasslands, such as less frequent mowing and woody- and exotic-species control, may replace some of the milkweed that has been lost from croplands.
Milkweed conservation has gained considerable interest because milkweeds are the host plants for monarch butterfly caterpillars. Public awareness of milkweeds is welcomed and has the potential to transform habitat conservation across midwestern North America. Scientists, we argue, can best aid milkweed and monarch conservation in two ways: (1) carefully considering the evidence for and against potential causes for monarch decline, with the most prominent being the connection between milkweed abundance and monarch populations, and (2) assessing the land-management practices that will best support milkweed and monarch populations in existing habitats and future restorations.
Monarch and milkweed population trends
Monarch butterflies (Danaus plexippus) migrate annually from a single Mexican overwintering locale in the state of Michoacán to broad regions across North America, east of the Rocky Mountains, in a migratory cycle that requires three to four generations of butterflies (Malcolm and Zalucki 1993). Mexican overwintering populations have declined precipitously since systematic censuses began in the mid-1990s, remaining well below the long-term average for the past decade (Brower et al. 2012). This negative trend has led to concern over the long-term viability of migratory monarch populations (Semmens et al. 2016), resulting in considerable efforts to conserve monarchs and welcomed public interest in their larval milkweed host plants (Borders and Lee-Mäder 2014, Jepsen et al. 2015). Nevertheless, conserving a species that uses multiple habitats and migrates over 2500 kilometers is a formidable task.
Currently, there are three nonmutually exclusive categories of hypotheses as to why monarch populations are declining. First, the loss or degradation of Mexican overwintering habitat may yield fewer initial migrants (Snook 1993). Second, mortality or stressors along the migration route due to pathogens, introduced inferior host plants, or habitat loss and degradation (e.g., diminished nectar sources, increased mortality traversing fragmented habitats, and nontarget insecticide effects) could reduce the survival or health of butterflies during the spring or autumn migrations (de Roode et al. 2008, Berenbaum 2015, Steffy 2015, Inamine et al. 2016). Finally, the loss of available milkweeds, the obligate larval host plants, across the summer breeding range to the north could limit oviposition success, caterpillar fitness, and/or adult recruitment (Pleasants and Oberhauser 2013).
The final hypothesis (the loss of host plants, known as the milkweed-limitation hypothesis) has gained particular attention, because it points to recent land-use and -management changes that could plausibly affect monarch butterflies by reducing their host populations (Pleasants and Oberhauser 2013, Pleasants 2015, 2016). Monarch caterpillars feed exclusively on milkweeds in the Asclepiadoideae subfamily of the family Apocynaceae, including true milkweeds in the genus Asclepias and their close relatives, such as Cynanchum laeve (figure 1; Yeargan and Allard 2005, Agrawal and Fishbein 2006). Isotopic and metabolite analyses have suggested that the most important host for monarchs in the summer breeding range are midwestern populations of common milkweed, A. syriaca (Malcolm and Zalucki 1993, Flockhart et al. 2013), whereas the initial spring migrants along the southern portion of the migration route tend to use earlier emerging milkweeds (e.g., A. viridis) that are more common there (Malcolm et al. 1992). Increased glyphosate herbicide use on resistant varieties of corn and soybeans (henceforth herbicide-tolerant, HT, crops) has coincided with a sharp decline in populations of A. syriaca within agricultural fields in the upper Midwest. Because milkweeds are highly susceptible to glyphosate herbicides, the connection between A. syriaca declines and glyphosate use is thought to be causal. At the same time, there is some evidence that milkweed populations within seminatural areas, such as Conservation Reserve Program (CRP) fields, have been fairly stable (Hartzler and Buhler 2000, Pleasants and Oberhauser 2013).

Swamp milkweed (Asclepias incarnata) is among monarchs most preferred hosts, and it was frequently encountered in our wetland surveys. However, because this habitat is comparatively uncommon across midwestern North America, swamp milkweed's overall importance for monarch caterpillars is less than that of other milkweeds.
The milkweed-limitation hypothesis has led to numerous important conservation efforts to bolster milkweeds, even though many scientists remain skeptical of the causal link between milkweed loss and monarch declines (Davis and Dyer 2015, Dyer and Forister 2016, Inamine et al. 2016). The skepticism of the loss-of-host-plant hypothesis largely revolves around the uncertainty in estimates of milkweed declines and monarch populations in the central portion of the monarch breeding range (the midwestern states of the United States). For milkweeds, surveys have only assessed one species, over a limited geographic range, in a limited number of habitat types (Pleasants and Oberhauser 2013). For monarchs, surveys across the breeding range have not shown the declines that would be expected if milkweed resources were limiting to the butterflies there (Badgett and Davis 2015, Howard and Davis 2015, Ries et al. 2015). But questions about these adult butterfly survey methodologies have led to uncertainty regarding their results (discussed below; Pleasants et al. 2016). It is not surprising, then, that demographic models using this uncertain data have pointed to different monarch life-cycle stages as being responsible for the declines observed on the wintering grounds (Flockhart et al. 2015, Inamine et al. 2016). This has in turn led to different conclusions regarding the key causal factors for declines: milkweed losses in the summer breeding range (Pleasants and Oberhauser 2013, Flockhart et al. 2015) or stressors or mortality outside of the summer breeding range, especially along the southward migration route (Inamine et al. 2016).
Adding data and assessing uncertainty in host-plant numbers
In this section, we use large-scale surveys across multiple habitat types to more comprehensively assess milkweed population trends in natural areas. We also compile available pre- and post-HT estimates of milkweed abundance in croplands (supplemental table S8). We then use this information, along with estimates of milkweed host-plant preference by monarchs and estimates of land-use change, to provide a comprehensive assessment of the change in monarch host-plant availability and abundance over the past two decades in Illinois. With each estimate, we highlight areas of uncertainty in data, calculations, and estimates. We also suggest ways to better evaluate the hypothesized links among HT crops, milkweed abundance, and monarch declines.
Have milkweeds declined in croplands?
The milkweed-limitation hypothesis originated in observations of milkweed loss from croplands due to increased glyphosate herbicide and HT crop use (Hartzler 2010, Pleasants and Oberhauser 2013). However, milkweed surveys in agricultural lands are generally quite rare, probably because milkweed is only a minor crop weed. Therefore, we compiled multiple sources of information on milkweed abundance before and after the adoption of HT crops (box 1; supplemental tables S1, S2). Although we find variation in milkweed estimates, the key finding is that each of these disparate data sources supported original conclusions and point to the same overall picture: a sharp decline in milkweed abundance in croplands (box 1). Studies in different midwestern states yielded decreases in milkweed abundance of between 93.7% and 96.5% in soy and corn croplands. The variation among estimates probably reflects differences in sampling methods, study sites, and the patchy distribution of milkweeds in fields, but the overall trend is clear (box 1). Supporting the causal role of glyphosates in these declines, milkweed abundance in two soy fields with a single glyphosate application declined by more than 70% over the season, whereas nonglyphosate treatments in both corn and soy had small to little effect on milkweed abundance (box 1; Pleasants 2015). Precision in estimates could be increased if future studies were to compare current milkweed abundances in HT crops with those in traditionally managed fields, thereby adding to the sparse available sample data (box 1).
Crop-cover accounts for a majority of land cover in Illinois and most midwestern states (Illinois State Geological Survey 2001). Two Asclepiadoid species (A. syriaca and C. laeve) occur in midwestern crop lands. Recent studies highlight the potential importance of A. syriaca in agricultural settings to monarch populations (Pleasants and Oberhauser 2013). We systematically reviewed the agricultural literature from Illinois to find estimates of A. syriaca as a weed in Illinois croplands before the widespread adoption of glyphosate-resistant crop types, and we compare these estimates with estimates from Iowa.
A. syriaca in Illinois croplands. No long-term studies recorded the long-term trends in milkweed abundance in Illinois croplands during the period of monarch decline. However, two agronomic studies provide snapshots of milkweed abundance in croplands prior to when glyphosate-resistant crop were common: Houghton (1973) found 173 milkweed stems per ha in corn and soy fields in 1965–1970, and Allen (1992) found 103 A. syriaca stems per ha in corn and soy in small plots in 1990–1991. In both cases, milkweed abundance was highly patchy with standard deviations of 145 stems per ha and 207 stems per ha, respectively (supplemental table S1). Differences in milkweed abundance between fields could only partially be explained by crop type and weed management practices. A downside of these studies is that they occurred in different fields with different nonglyphosate practices. Post-glyphosate A. syriaca abundance in croplands was reported by Schutte and colleagues (2010) in a study of glyphosate retreatments. They found 8.51 A. syriaca stems per ha in 133 glyphosate-treated fields in 2005–2006 (standard deviation = 38.7 stems per ha) and no C. laeve. Estimates from these studies suggest a 93.7% decrease in milkweed abundance in croplands from pre- to post-glyphosate treatments.
A. syriaca in Iowa croplands. Two surveys tracked milkweed abundance in croplands from the late 1990s to the late 2000s. Hartzler (2010) surveyed milkweed patches in crop fields in 1999 and 2009 and found a sixfold decrease in the number of milkweed patches between those surveys and a 96.5% decline in estimated abundance. This study used standard survey practices; however, the study also focused on milkweed patches relatively close to (less than 100 meters from) roads, so extrapolation from these studies to a whole field estimate that is needed to calculate total crop area across the region may not be possible. Pleasants and Oberhauser (2013) repeatedly surveyed milkweed patches in fields starting in 2000. By 2008, they found a virtual extirpation of the 998 patches they had initially surveyed. Pleasants (2015) reported a sharp (more than 70%) decrease in A. syriaca abundance over a single growing season in soy fields with glyphosate use but a small or no decrease following nonglyphosate herbicide treatments.
A. syriaca in other croplands. Yenish (1997) reported 100–5000 stems per ha of A. syriaca in cornfields and 10000–35000 A. syriaca stems per ha in soy fields in Maryland in 1990–1991, before glyphosate resistant crop adoption. A. syriaca was present in both conventional and no-till fields that lacked post-emergence glyphosate treatment in Ontario in 1994–1996 (Swanton et al. 1999). Both of these estimates are greater than baseline Corn Belt estimates of milkweeds from Illinois and Iowa.
Summary and needed evidence. Evidence supports a sharp decline in A. syriaca abundance in croplands at Iowan and Illinoisan sites. High-quality information on agricultural milkweed trends for the rest of the midwestern region is lacking, but the literature supports a patchy milkweed presence in crops prior to glyphosate-resistant crops. Milkweeds are susceptible to glyphosate herbicides (Pline et al. 2000), and glyphosate application is intended to reduce populations of weeds such as milkweed. Large-scale studies of present-day herbicide, tillage, and other crop management practices on milkweed abundance are needed. These studies may identify weed-management practices that control economically damaging weeds but lack strong impacts on minor weeds such as milkweed and better explain how monarchs use agricultural milkweeds across a range of densities.
Milkweed surveys in natural areas
The milkweed-limitation hypothesis pointed to agricultural milkweeds as the causal factor for monarch declines (Pleasants and Oberhauser 2013). At the same time, in more natural areas, the estimates of abundance have been of limited scope with respect to the breadth of habitats and the milkweed species investigated (Hartzler 2010, Pleasants and Oberhauser 2013). Therefore, it is difficult to assess whether milkweed populations in nonagricultural lands have been stable or are declining across the vast summer breeding range, such that they might also be contributing to monarch losses or nullifying milkweed declines in agricultural areas. We used a long-term botanical survey covering the predominant natural and seminatural vegetation types across Illinois to estimate milkweed trends. The survey used randomly selected sites from grassland, wetland, and forest habitats (198, 187, and 160 sites, respectively), which it tracked every 5 years since 1997 using standardized plant sample plots (Carroll et al. 2002; see supplemental information for further survey description). We pared this sample to the 85 grassland, 49 wetland, and 7 forest sites where milkweeds were observed in sample plots (figure 2) to determine changes in milkweed abundance over time (Spyreas et al. 2016). Because this study includes randomly selected sites and because of the similarity in vegetation patterns and land cover between Illinois and the rest of the midwestern United States’ “Corn Belt” region (Taylor and Shields 2000), trends here can be extrapolated across this most productive portion of the monarch butterfly's summer range (Wassenaar and Hobson 1998).
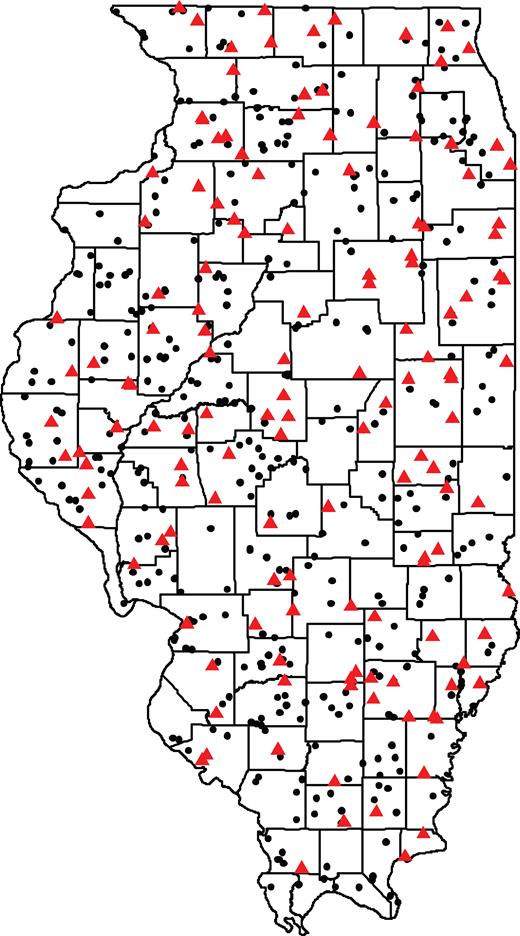
Grassland, wetland, and forest survey sites across the state of Illinois. The red triangles represent the survey sites with at least one of the target species included in our analyses. The black circles represent sites where our target species did not occur.
We found that the average milkweed species abundance varied considerably among sites and habitats, with the four most common species being A. syriaca, A. verticillata, C. laeve, and A. incarnata (figure 3, supplemental table S3, supplemental figures S1, S2). We then analyzed long-term trends in the abundance (log [cover+1]) of these four most common milkweed in their key habitats using a mixed-model approach. These four species were chosen because they were common enough for formal statistical analysis. We also looked for site-level correlates with the abundance of these species using permutation tests at sites that were visited multiple times (48 grassland sites for A. syriaca, 17 grassland sites for A. verticillata, 16 grassland sites for C. laeve, and 25 wetland sites for A. incarnata; supplemental table S4). Forests were not included in these statistical tests of milkweed abundance trends because of the small sample size. The details of the survey and statistical methods can be found in the supplemental information.
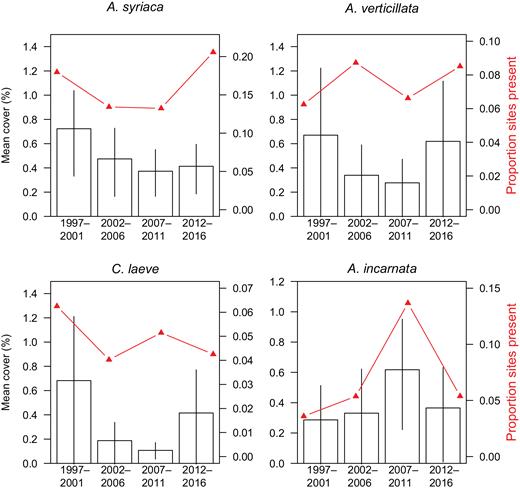
The abundance trends for the most common (proportion of sites present) and abundant (mean cover per quadrat) milkweed species in the habitat that they most frequently occurred. The error bars represent 95% confidence intervals, bootstrapped with 99,999 resamples. None of the species abundances showed statistically significant change through time (in all cases p > .15)
Several lines of evidence fail to show milkweed declines in natural areas across the Midwest. Our survey shows that none of the species we tested are declining in their natural and seminatural habitats (figure 3). Despite apparent declines in A. syriaca and C. laeve abundance in grasslands (figure 3a, 3c), the linear mixed-effect models did not find these trends to be significant (A. syriaca, ΔAIC = –5.66, χ2(1) = 1.9, p = .16; C. laeve, ΔAIC = –5.73, χ2(1) = 1.01, p = .31). Temporal trends in A. incarnata in wetlands and A. verticillata in grasslands were also not significant (A. incarnata, ΔAIC = –6.45, χ2(1) = 0.30, p = .58; A. verticillata, ΔAIC = –6.92, χ2(1) = 0.09, p = .76).
With regards to habitats, we found that grasslands harbor greater milkweed abundance per site than other habitats do (table S3, figure S2), and they account for more land cover in Illinois (13%–16%), meaning that they contain a far greater overall proportion of the state's milkweeds than other habitats do. Wetlands, by comparison, harbor high densities of milkweeds, but they constitute relatively little land cover (0.45% of Illinois's area; supplemental tables S4, S5). Forests are common across the state (table S5), but they contain far fewer milkweed plants (table S3).
In order to relate milkweed abundance trends to the resources available to monarchs, we ranked each milkweed species for their relative value as monarch caterpillar hosts (termed the coefficient of relative host use [HU]). We used five studies that observed and quantified the relative abundance of naturally occurring caterpillars on wild host plants (Price and Willson 1979, Chaplin and Walker 1982, Betz et al. 1997, Bartholomew and Yeargan 2001, Roels 2011, supplemental table S6). We then estimated the change in (food) resources available to monarch caterpillars across Illinois by translating each milkweed's abundance into monarch resource units (MRU). MRU values for natural areas were calculated as the product of (a) the mean percent cover for a milkweed species in a given habitat, (b) the percentage of Illinois's area covered by that habitat, and (c) the coefficient of relative host use (HU) of that milkweed species (figure 4). The MRU values were summed across species for each 5-year period of the study. We found multiple estimates of the area in Illinois covered by grassland habitat and how that area changed over the study period (supplemental table S7). In order to incorporate the uncertainty associated with this parameter, we created three possible scenarios for change of MRU through time: a best-estimate scenario, an optimistic scenario, and a pessimistic scenario (supplemental information, supplemental tables S7, S8, S9, S10).
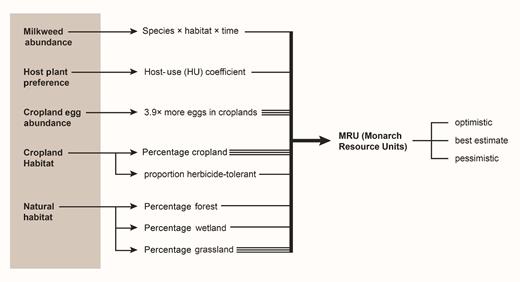
Information that was incorporated into estimates of monarch resource units (MRU). The compilation of information into MRU was conducted using equation S1 in the supplemental information. The triple lines indicate parameters whose uncertainty yielded three different MRU estimates.
The region-wide estimate of monarch host-plant resources measured as MRU declined by 36% (between 33%–42%) over the 20-year study period in natural areas (figure 5). These trends were largely driven by trends in A. syriaca (and to a lesser extent C. laeve), as well as the decrease in area covered by grasslands. Alternatively, A. verticillata is relatively common in grasslands, but because it is a relatively poor monarch host, it contributed little to MRUs. Similarly, A. incarnata is a high-quality monarch host, but the emergent wetlands where it occurs cover a small portion of Illinois (table S5). Therefore, we report a decline in the abundance of milkweed resources for monarch larvae, although there was no statistically significant decline in any single milkweed species we tested.
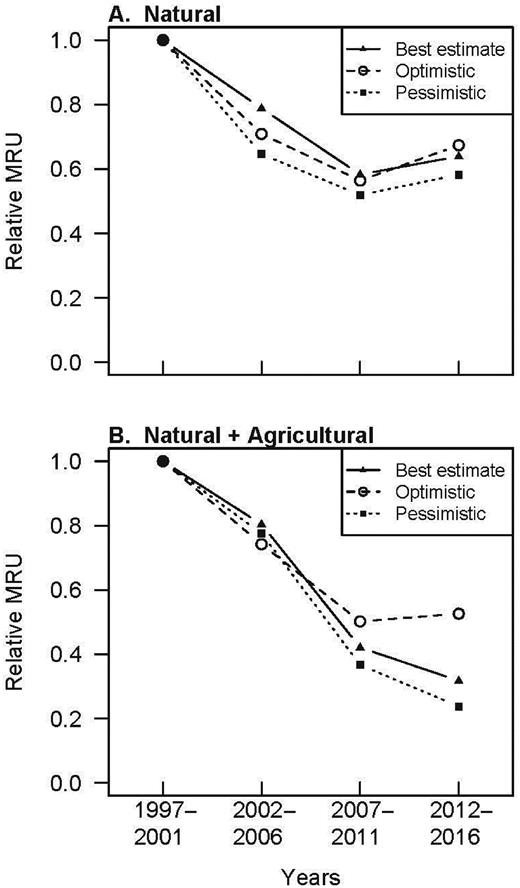
Trends in monarch resource units (MRU) across Illinois from 1997 to 2016. MRU reflects available monarch host resources. Trends in MRU are shown for Illinois (a) grasslands, wetlands, and forests (top panel; CTAP sites only) and (b) from the combination of these natural habitat estimates, as well as Illinois cropland estimates (bottom panel). We attempted to capture uncertainty in parameter estimates by representing three different possible scenarios, which include the bounds of our MRU estimate and our best estimate.
Past milkweed surveys on nonagricultural lands are not easily extrapolated regionally because of their sample designs; however, they also tend to show no or small milkweed declines across natural areas (Hartzler 2010, Pleasants and Oberhauser 2013). Hartzler (2010) did not observe a decline in the number of A. syriaca patches along roadsides in Iowa between 1999 and 2009. Using data from volunteers from the Monarch Larval Monitoring Project, Pleasants and Oberhauser (2013) reported a 31% decrease in area of plots in nonagricultural habitat covered by milkweed patches.
Combining milkweed estimates across habitats to assess host-plant losses across the entire region
Milkweeds host monarchs across multiple habitats and land-use types, and it is the combined effect of all habitats that is relevant for monarch populations. We combined the MRU trends from natural areas with those from croplands to assess total MRU across Illinois (assuming that all agricultural MRU would be contributed by A. syriaca; figure 4, supplemental equation S1, supplemental methods). We estimated the relative land area of croplands and grasslands on the basis of yearly estimates of land cover from the National Agricultural Statistics Service Cropland Data Layers (USDA 2015). MRU from agricultural areas incorporated (a) the percentage of the state planted in the two main row crops of the region, corn, and soybeans (table S8); (b) the proportion of each crop type that was genetically modified to be herbicide tolerant (table S8); (c) the densities of common milkweed in fields with and without herbicide-tolerant crops (table S9); (d) the HU value for common milkweed (table S6); (e) the area covered by a single stem of common milkweed (used to convert between stems per hectare (ha) from agricultural studies to the percent cover from CTAP data; table S10); and (f) the ratio of monarch use as a larval host of agricultural milkweeds and nonagricultural milkweeds (figure 4, equation S1; Pleasants and Oberhauser 2013). To capture the variability in parameter estimates, we again used three scenarios for our estimate of MRU. The parameters that varied between scenarios were the densities of milkweed in crop fields, the area covered by a single milkweed stem, and the ratio of monarch use of agricultural versus nonagricultural milkweeds. In every scenario, total MRU declined (figure 4). Our best estimate is that MRU declined by 68% across the state over the last 20 years. The upper end of our plausible range of MRU decline (47%–76%) is similar to but less than the decline in larval production calculated by Pleasants and Oberhauser (2013) over a similar time period and less than the updated estimates from Pleasants (2016).
Another key outcome of these analyses was finding that three parameter estimates stood out as being by far the most influential for MRU estimates. The first parameter is how much area is covered by grassland habitat in Illinois and how much that has changed, especially since the widespread adoption of corn ethanol as a gasoline additive (Lark et al 2015). Second, our scenarios differ with respect to relative demographics of monarch reproduction on agricultural versus nonagricultural milkweeds. Multiple authors report the increased reproductive output associated with milkweeds in croplands (Bartholomew and Yeargan 2001, Pleasants and Oberhauser 2013), but those estimates do not investigate how relative success changes spatially and along milkweed-density gradients (Flockhart et al. 2012). This increased cropland output effect (increased Cropland Egg Abundance in figure 4) is the single most crucial parameter to monarch host trends, adding a 13% MRU loss to estimates (figure 5b) and tripling the amount of milkweed restoration that would be needed to counter loss from croplands (see “How do we manage milkweed populations?”). Finally, the estimated milkweed densities in croplands pre- and post-HT crops are also moderately influential for MRU values. Variation arises here because historical study methodologies were conceived for other purposes, making this source of uncertainty unavoidable. Going forward, it would behoove us to obtain more accurate estimates of these three parameters, because refining them could dictate conclusions regarding this system and ultimately guide future decisions regarding monarch conservation.
In summary, the best available data indicate that milkweeds have declined precipitously in croplands. In natural areas, declines are more moderate and declines were not statistically significant for any single species. Steep cropland declines have now been shown multiple times across multiple states (box 1; Pleasants and Oberhauser 2013). Even so, we now have far more robust surveys and trend data for natural areas’ milkweed populations. Therefore, if monarchs are declining because of overall losses in milkweed abundance, then that decline must implicate croplands and the increased adoption of HT crops. However, we must caution that the estimates of adult monarch numbers in Illinois—or the breeding range generally (Ries et al. 2015)—do not closely follow the long-term trends observed for MRUs. We discuss this discrepancy in detail in the following section.
From milkweeds to monarchs: What we know, what we don’t know, and how to move forward
The monarch life cycle is complex, and monarchs face numerous challenges throughout their life cycle and across their range, declines in milkweed hosts being only one of them. To assess the role that each of these challenges play in overall population trends, ecologists construct demographic models that combine an organism's numbers at different life stages. In two recent models of monarch butterfly demography, Flockhart and colleagues (2015) and Inamine and colleagues (2016) asked which part of the monarch life cycle and migratory journey have contributed the most to monarch declines. Their results differed in their support for the milkweed-limitation hypothesis, so it is worthwhile to compare these models for the constitutive uncertainties in their estimates. The key difference between the two models is in the importance that each model gives to adult butterfly censuses in the breeding range versus estimates of larval and egg production from the breeding range—the latter being inextricably tied to estimates of milkweed abundance.
Flockhart and colleagues (2015) created an impressive demographic model that included both mature and immature life stages and incorporated the entire migration route as four discrete sections. In the wintering range, they parameterized their demographic model with estimates of adult monarch abundance in Michoacán, Mexico. In the breeding range, they used estimates of survival and fecundity for immature and mature life stages, as well as estimates of milkweed-abundance trends in a variety of habitats. Flockhart and colleagues (2015) found that monarch population growth was most sensitive to larval life-stage dynamics (especially in the central portion of the breeding range, where most of our study area occurs), but population trends were also quite sensitive to adult dynamics, especially in the southern portion of the breeding range. The model proposed by Flockhart and colleagues (2015) did not include estimates of adult abundances in the breeding range that fail to show a decline (Badgett and Davis 2015, Howard and Davis 2015, Ries et al. 2015). As a result, the model was highly dependent on accurate estimates of milkweed abundance and vital rates in the breeding range, where any inaccuracy in these parameters results in high uncertainty in adult breeding-range numbers (Inamine et al. 2016). Our estimate of the proportion of milkweed abundance apportioned to agricultural areas is substantially smaller than the assumptions of Flockhart and colleagues (2015). We found 22% of all milkweed abundance (26% when weighting by the host-use coefficient, 35% when only considering common milkweed) was in agricultural areas during the 2012–2016 period (figure S2), versus the estimate by Flockhart and colleagues (2015) of 67% in 2013. Flockhart and colleagues (2015) covered a larger area than our study, although the area of agricultural land in Illinois is larger than the whole of their larger study area. This discrepancy is likely influential in determining the most important life stages and geographical regions for monarchs, and our results may inform a key point in this valuable demographic model.
Inamine and colleagues (2016) estimated monarch demography on the basis of adult surveys across the migratory range (Brower et al. 2012, Badgett and Davis 2015, Howard and Davis 2015, Ries et al. 2015). They demonstrated strong, positive correlations in monarch abundances among different portions of the northward migration from Mexico to the Midwest, but they found a poor correlation between summer breeding-range adult estimates and subsequent autumn and overwintering population estimates. The authors suggested that high and variable mortality during southern migration caused the poor correlation. Their model relied heavily on estimates of adult butterflies, which are surveyed in fairly consistent ways (Ries and Oberhauser 2015). The key source of uncertainty in this model is the degree to which adult surveys within the breeding range represent total butterfly production and numbers. Pleasants and colleagues (2016) argued that these adult breeding range surveys fail to show a monarch decline because of survey artifacts, including inaccurate sampling and a bias toward sampling nonagricultural habitats, where monarch losses are not expected. In response to this criticism, Dyer and Forister (2016) agreed that some of the adult survey numbers could be artifactual, but barring empirical evidence showing inadequacy in the survey techniques or a decoupling of monarch populations in natural and agricultural habitats, the results from these massive data sources must not be disregarded. In addition, it is not clear why survey artifacts would reduce the ability to detect correlations in one direction (northern migration) but not the other (southern migration). The Inamine and colleagues (2016) model suggests that monarch mortality has increased in the southern range over time. At present, it is not clear which factor or factors in that region coincide with the timing of monarch loss, and finding such factors is crucial to establishing what measures could be taken to prevent monarch loss during the southward migration.
The results of Flockhart and colleagues (2015) and Inamine and colleagues (2016) both suggest that monarch population dynamics can be highly influenced by adults in the southern portion of the breeding range, but they disagree starkly on the importance of larval dynamics farther north (the epicenter of the milkweed-limitation hypothesis). Both models’ main criticism has been uncertainty in their data (especially milkweed abundance, larval vital rates, and adult census numbers). We highlight key expectations that can be used to distinguish the assumptions of the two models.
Flockhart and colleagues (2015) included density-dependent larval mortality as an explanatory factor in monarch declines on the basis of observed patterns in a manipulative study (Flockhart et al. 2012); monarch larvae experience higher mortality because of higher densities per milkweed. Under the Flockhart and colleagues (2015) model, we might expect greater mortality of larvae in years with greater numbers of colonists and smaller numbers of milkweeds. One concern in this approach is that the densities of monarch eggs per plant that resulted in observed density-dependent mortality (Flockhart et al. 2012) were two to five times higher than the numbers of eggs observed in natural lands where monarchs would be expected to aggregate in the absence of cropland milkweeds (Oberhauser et al. 2001). This disparity may be inconsequential, however, because density dependence could enter the model at other points, such as if adult monarchs spend extra time searching for unoccupied milkweeds and die with a greater proportion of unlaid egg loads when milkweeds decline. Thus, the increasing mortality of larvae or increasing unlaid egg loads are clear and testable expectations of the Flockhart and Colleagues (2015) model.
One expectation from the Inamine and colleagues (2016) model is that monarch population growth between overwintering and breeding zones must be positive, in order for the decreasing number of colonists over time from the overwintering population to result in stable adult numbers in the breeding range. This may have necessitated an increasing reliance on nonagricultural milkweeds over time and an increase in natural area larval and egg densities per plant (assuming monarch population growth is sensitive to larval dynamics; Flockhart et al. 2015). This expectation does not match larval census data that have failed to show an increase of oviposition on milkweeds in natural areas (Stenoien et al. 2015). The Inamine and colleagues (2016) model does not expect increasing density-dependent mortality or increased host search times (and proportions of unlaid eggs) in the breeding range to account for monarch declines. Surveys of monarch larvae do show increasing larval mortality in the breeding range over time (Nail et al. 2016), although the timing of this increase is not entirely consistent with either the Inamine and colleagues (2016) or the Flockhart Colleagues (2015) models.
To confidently assess the role of milkweed loss in monarch declines, researchers will need to fill in the aforementioned gaps in current modeling studies. This will involve a detailed examination of arcane aspects of butterfly surveys, expansion of monarch life tables, and plausible mortality factors over the entire range, and this will take time and resources. At present, we show clear losses of milkweed in the Midwest, although not quite to the extent of some evidence that has been used in support of the milkweed-limitation hypothesis. Nonetheless, we conclude that milkweed loss in the northern breeding range and stressors on adults in the southern portion of the breeding range (most likely during autumn migration) remain the most compelling agents of monarch declines. Multiple simultaneous stressors were implicated in previous extinctions of once-abundant, migratory North American animals, such as the passenger pigeon (Hung et al. 2014). We argue that both potential stressors should be considered as potentially important factors in monarch declines. Despite uncertainty and the likelihood of multiple causes of decline, milkweed restoration remains one immediately actionable strategy to combat monarch loss.
How do we manage milkweed populations?
Because we do not know whether we have reached a threshold in milkweed host-plant resources that could be contributing to monarch declines, there have been recent attempts to boost milkweed numbers via plantings and better habitat management. Although our data indicate that milkweed losses from natural and seminatural areas are unlikely to contribute to monarch declines, the establishment of greater milkweed populations has many benefits and could help to counter losses from agricultural lands. Our data suggest that milkweed restoration could replace cropland losses if the “optimistic” scenario is accurate (figure 5). In this case, an increase of common milkweed abundance by 84% in natural areas—by either increasing their abundance to 0.24% cover in existing natural areas (approximately 380 stems per ha) or by adding 1,700,000 ha of natural grassland—would counter agricultural losses. However, these estimates do not consider negative density dependence (Flockhart et al. 2012), and an increase in the number of milkweeds is unlikely to result in exactly the same proportional increase in monarch population size. Under the best-estimate scenario, an increase of milkweed abundance by 670% in natural areas would be needed to counter agricultural losses (pessimistic: 2250%)—requiring that the entire area of Illinois be converted to grasslands (figure 6). Moreover, the impacts of dramatically increased milkweed abundance on native biodiversity are unknown and possibly detrimental, because milkweed was likely never at such high densities in midwestern grasslands (Pleasants 2015). Even if fully compensating for agricultural losses is not possible or achieved, the simple management practices highlighted below (box 2) would boost milkweed abundances.
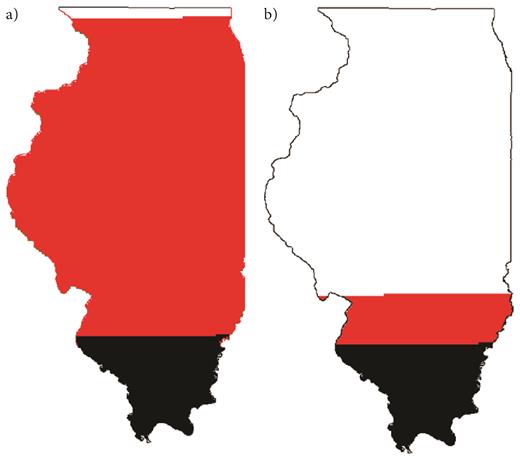
New grassland plantings needed to compensate for milkweeds lost from Illinois croplands under (a) the best estimate and (b) optimistic milkweed-loss scenarios. The black portion of the state represents our estimate for current grassland cover (supplemental table S7). The red portion represents the area of additional grassland cover that would be needed to compensate for milkweed losses since the adoption of herbicide-tolerant crop technology (assuming the same milkweed abundance in planted versus existing grasslands).
Site managers are tasked with restoring habitat for milkweeds as monarch hosts. They need to know what site characteristics and management promote milkweeds. From an analysis of change in milkweed abundance across randomly chosen sites in Illinois, we describe site-level characteristics and management practices that correlate with increasing milkweed abundance for the four most abundant midwestern milkweeds in their key habitat types.
A. syriaca (grasslands)
Sites with little to no trees, that are not too shrubby, and where overall vegetation cover did not increase too much. Sites where plant diversity was moderate and where plant diversity increased over time (figure 5, supplemental table S11).
A. verticillata (grasslands)
Sites that were mowed frequently. Sites near croplands. Sites with disturbance-associated plant communities with many exotic species but that were not being invaded by additional exotics (supplemental table S12). Sites with lower diversity.
C. laeve (grasslands)
Sites surrounded by croplands and not surrounded by forests. Sites heavily invaded by exotic plant species and with low floristic quality. Sites that were mowed or disturbed frequently. Degraded sites whose plant diversity was decreasing (supplemental table S13).
A. incarnata (wetlands)
Sites with fewer nonnative species, sites that had less nonnative cover and that were not increasing in nonnative cover. Sites that were not decreasing in overall vegetation cover. Sites with higher plant diversity (supplemental table S14).
In order to successfully manage for milkweeds, it is crucially important to understand the factors that affect long-term milkweed abundance. Although the climate that defines the geographic range of milkweeds is well known (Lemoine 2015), it is far less clear what site-level factors correspond to stable or increasing milkweed abundance. These factors are particularly important because local factors such as mowing, exotic plant invasions, or buffers around natural areas can be easily manipulated by land managers (Borders and Lee-Mäder 2014, Fischer et al. 2015).
Each milkweed species’ trend in the Illinois survey correlated with a different set of site variables (box 2; supplemental tables S11–S14). A large number of correlates were tested for each species (44 statistical tests), and every species had a greater number of statistically significant results than would be expected randomly at the α = 0.05 level (each species had at least nine significant results, two-sided exact binomial test p < .0003). These variables were largely consistent with descriptions of their preferred habitats in regional floras, suggesting that management practices and targets for optimal milkweed habitats for the different species are readily identifiable. For example, site-level correlates of population increases in A. verticillata and C. laeve suggested that disturbed, early-successional sites are optimal for these milkweeds, which is not surprising given that they are abundant along roads and grassland rights of way. Management for these species is not needed because such habitats are ubiquitous. Alternatively, we found that A. syriaca abundance increased the most in grasslands with increasing vascular plant diversity (figure 7), where the number of nonnative species was decreasing, and sites that did not have too much woody vegetation. This is consistent with A. syriaca as a plant species that thrives in old-field habitats and midsuccessional grasslands, which would not tolerate frequent severe disturbance, but in the absence of occasional, moderate disturbance or ecological management will quickly be excluded as the habitat transitions to shrubland or forest in the Midwest (Bazzaz 1975). It also suggests that sites whose land management promotes low diversity (such as with frequent mowing, herbiciding, overgrazing, or species-poor plantings) result in poor A. syriaca habitat. Attention should also be paid to the timing of disturbance in grasslands, because mowing during monarch larvae maturation is suggested as a key cause of monarch mortality by creating grassland population sinks (Fischer et al. 2015). At a larger scale, promoting A. syriaca would rely on restoring a dynamic grassland landscape mosaic with grassland and wetland habitats at all successional stages.
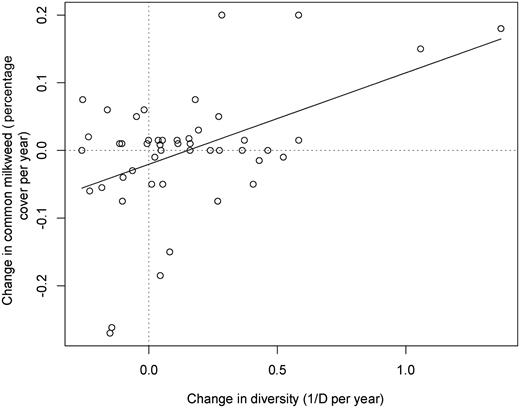
The correlation between changes in common milkweed abundance and changes in plant diversity at the site level. The inverse Simpson's Index (1/D) was used to quantify diversity, and the percent cover was used to quantify milkweed abundance. The rate of change in both the explanatory and response variables was calculated as the change in the value divided by the number of years between the first and last site visits: r = .474, found to be significantly greater than 0 after bootstrapping with 99,999 resamples (supplemental table S11).
Management to promote milkweeds and diverse grasslands will have numerous ancillary benefits beyond its potential to bolster monarch numbers. Several milkweed species that can support monarch butterflies are themselves rare or endangered (Roels 2011, Kim et al. 2015). Milkweeds are host to a diverse and charismatic specialist insect community, including Tetraopes and Chrysochus beetles (Van Zandt and Agrawal 2004). Milkweeds and their associated grassland forbs provide important nectar resources, whose declines have important ramifications for grassland insects generally, in addition to monarchs along their migration route (Tooker et al. 2002).
What about rare or less-palatable milkweed species?
Currently the only 3 abundant milkweed species that are effective monarch hosts in Illinois are A. syriaca, A. incarnata, and C. laeve, although 19 milkweed species are native to the state (Pearse et al. 2016). The rest are either rarely selected by ovipositing adults or are too unpalatable or rare to substantially contribute to monarch populations. Have any of these species been more important monarch hosts in the past, and could they be in the future? Historic accounts indicate that A. syriaca has long been a common component of the midwestern flora, but other milkweed species were certainly far more common before European settlement and the plowing of the prairies (Short 1845, Gray et al. 1889, Pleasants 2015). Several milkweed species in Illinois are suitable monarch hosts (Erickson 1973; table S6), but most are so rare as to not contribute substantially to MRUs. A. tuberosa is an increasingly popular component of prairie restorations and urban prairie plantings, so its monarch importance may increase. The suitability of several other milkweed species as monarch hosts is unknown. Finally, given that the range of C. laeve has expanded northward over the past 50 years and that A. syriaca has expanded its range south (Malcom et al. 1992, Pearse et al. 2016), assessments of the most important larval milkweed food species for producing adults may need revisiting, because they were established 20 or more years ago (Wassenaar and Hobson 1998).
What about unpalatable milkweeds? Populations of other Lepidopteran species have been known to dramatically increase after expanding their diet to include other common plant hosts. Monarch butterflies themselves have expanded their geographic range into the Eastern Hemisphere in association with the novel colonizing Asclepiadoid host species Gomphocarpus fruticola and G. physocarpus (Fernández-Haeger et al. 2015). It is unclear whether the host expansion of monarchs onto Gomphocarpus species involved evolutionary processes on the part of the butterfly or whether those species were already ecological fits to the historical monarch diet (Agosta 2006). Such a host expansion may be less likely for monarchs because of their low genetic diversity, but the potential for expanded host-plant use seems plausible given that preferred host use for them switches dramatically among milkweed species seasonally and regionally across the monarch's migration. As such, it would be worthwhile to track monarch use across milkweed species and habitats that are currently assumed to be less used or productive for monarchs.
Conclusions
Populations of milkweed species in midwestern natural areas have been relatively stable over the past 20 years, during which time cropland numbers have declined. If monarch declines are linked to milkweed losses in the Midwest, they must be associated with cropland losses as opposed to more natural habitats. Modeling approaches disagree on the likelihood of the milkweed loss–monarch decline connection, and a focus on improving estimates of larval vital rates and adult breeding-range surveys will be important to rectify differences in those models. Management practices that promote diverse, native grasslands and wetlands at a variety of succession states will improve habitat for A. syriaca, which may be important for monarchs because this is the largest component of monarch host-plant resources in the Midwest.
Acknowledgments
We thank the many CTAP scientists and crews who collected the data used here, especially James Ellis, Connie Carroll-Cunningham, and Tim Rye. Support for the CTAP program comes from the Illinois Department of Natural Resources. Adam Davis, Brian J. Schutte, and Aaron Hager very generously allowed us the use of their Illinois cropland weed data. Anurag Agrawal, Tari Tweddale, John Pleasants, May Berenbaum, Karen Zaya, Danielle M. Ruffatto, and the three anonymous reviewers made valuable contributions to the project. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government. The data and analyses for the study were archived on the University of Illinois IDEALS archive at doi:10.13012/B2IDB-5072223_V1.
Supplemental material
Supplementary data are available at BIOSCI online.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}