



Abstract
The development of social emotions such as compassion is crucial for successful social interactions as well as for the maintenance of mental and physical health, especially when confronted with distressing life events. Yet, the neural mechanisms supporting the training of these emotions are poorly understood. To study affective plasticity in healthy adults, we measured functional neural and subjective responses to witnessing the distress of others in a newly developed task (Socio-affective Video Task). Participants’ initial empathic responses to the task were accompanied by negative affect and activations in the anterior insula and anterior medial cingulate cortex—a core neural network underlying empathy for pain. Whereas participants reacted with negative affect before training, compassion training increased positive affective experiences, even in response to witnessing others in distress. On the neural level, we observed that, compared with a memory control group, compassion training elicited activity in a neural network including the medial orbitofrontal cortex, putamen, pallidum, and ventral tegmental area—brain regions previously associated with positive affect and affiliation. Taken together, these findings suggest that the deliberate cultivation of compassion offers a new coping strategy that fosters positive affect even when confronted with the distress of others.
Introduction
Compassion can be defined as “the emotion one experiences when feeling concern for another's suffering and desiring to enhance that individual's welfare” (Keltner and Goetz 2007). In other words, compassion is composed of an other-related affective component and a prosocial motivational component. In the context of the present study, we investigated compassion as rooted in a contemplative tradition aimed at cultivating feelings of warmth, love, and care toward a sequence of people in a state of quiet concentration (Fredrickson et al. 2008; Leiberg et al. 2011). More specifically, the training focused on the practice of loving kindness (“Metta” in Pali) that aims to foster an attitude of emotional positivity, benevolence, and friendliness, as it is naturally experienced toward close loved persons like one's own mother or children (similar to strengthening modes of affiliation). During the training, this attitude was extended toward all persons independent of context and preferences.
To reliably investigate how cultivating feelings of warmth and prosocial motivation influences neural responses when exposed to the suffering of others, we developed a new paradigm—the socio-affective video task (SoVT)—in which participants saw short video sequences based on real-life situations depicting others in distress. Experiencing feelings of loving kindness toward another person's affliction is best described with the term “compassion.” In other words, compassion can be regarded as applying loving kindness (as trained) in response to the suffering of others (implemented by the SoVT). Thus, although the concepts of compassion and loving kindness are separable as the object they refer to can differ, we will for reasons of simplicity and clarity generally refer to compassion and compassion training in the present paper.
Even though closely related, compassion (feeling for someone) differs from empathy, which is defined as feeling with someone, that is, sharing the other person's emotion (de Vignemont and Singer 2006; Batson 2009; Singer and Lamm 2009). As such, empathic responses to witnessing another person's suffering usually involve shared negative affective experiences. Indeed, most neuroscientific studies on empathy to date have focused on the domain of vicarious pain. These studies revealed that a network including the anterior insula (AI) and the anterior medial cingulate cortex (aMCC) is involved both in the first-hand experience and the vicarious experience of pain (Fan et al. 2011; Lamm et al. 2011). Furthermore, activity in aMCC has been repeatedly shown to be modulated by the experience of unpleasantness (Rainville 2002). Despite these advances in empathy research, the neural correlates underlying compassionate feelings are still poorly understood. Furthermore, it has remained unresolved which neural processes are relevant for strengthening such prosocial emotions.
Although neural plasticity has been studied for decades (Pascual-Leone et al. 2005), the majority of research has favored the investigation of neural changes induced by the training of motor and cognitive abilities (Karni et al. 1995; Olesen et al. 2004), while the study of neural plasticity underpinning socio-affective competences has long been neglected. To date, research in the domain of affective plasticity has focused mainly on short-term changes associated with fear conditioning and fear extinction (Gottfried and Dolan 2004; Schiller et al. 2008). The ability to cope with difficult situations is crucial for successfully maneuvering through our social lives, as well as for maintaining mental and physical well-being (Gross and Muñoz 1995). Therefore, the lack of studies focusing on the neural mechanisms underlying the plasticity of positive social affect is surprising. The neuroscience of compassion is actually still in its infancy and has, so far, only been studied cross-sectionally (Lutz et al. 2008; Beauregard et al. 2009; Immordino-Yang et al. 2009; Kim et al. 2009; Simon-Thomas et al. 2011). To fill this gap, we investigated functional neural plasticity induced by compassion training in healthy adults in a series of functional magnetic resonance imaging (fMRI) experiments that combine multiple approaches with the aim of obtaining measures that are robust in terms of replicability, validity, and generalizability (Fig. 1A and Supplementary Fig. S2).
{kind=link}
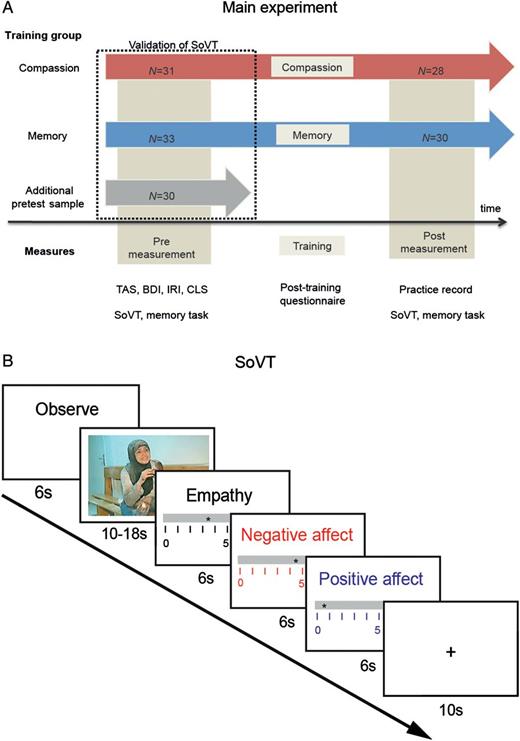
Experimental design. (A) Main experiment. At pre-training measurement, a total of 94 women were scanned by means of fMRI while viewing 1 of 3 parallel video sets from the newly developed SoVT. Thirty of 33 participants completed the memory training, 28 of 31 participants completed the compassion training, and 30 participants were included as an additional validation sample at pre-training measurement. Pre-training measurement included the trait questionnaires Toronto Alexithymia Scale (TAS, Bagby et al. 1994), Beck's Depression Inventory (BDI, Beck et al. 1996), IRI (Davis 1983), and CLS (Sprecher and Fehr 2005), as well as the memory task and fMRI measurements in response to the SoVT. After the training day, self-report data about the training were collected in form of a post-training questionnaire. At post-training measurement, participants handed in their practice records, performed the memory task and were scanned while viewing the SoVT. (B) SoVT. In the main experiment, each fMRI measurement presented participants with 12 HE and 12 LE videos (duration 10–18 s). After each video, participants rated their subjective experience of empathy, positive affect, and negative affect. Every mini-block of 3 HE or LE videos was followed by a fixation cross (displayed for 10 s).
First, we assessed the neural networks underlying the deliberate generation of compassion in an expert long-term meditation practitioner immersed in different types of compassionate states. Subsequently, we carried out 2 compassion training experiments in meditation-naive participants (total N = 46) from which we derived independent regions of interest (ROIs). These findings were used to qualify the neural changes observed in a third longitudinal experiment which compared compassion training in 28 female participants with an active control group (N = 30) who received training in a memory-enhancing technique (Bower 1970). To reliably assess participants' compassionate responses, we specifically developed the SoVT, which measures empathy, positive affect, and negative affect in response to witnessing videos depicting people in distressing situations (high emotion videos, HE) or showing people performing everyday activities (low emotion videos, LE). The task was composed of 3 parallel video sets which can be used interchangeably to avoid habituation induced through a repeated measurement, thus making it optimal for longitudinal designs.
In keeping with previous findings on the neural substrates of empathy, we expected that empathy for HE videos would be accompanied by activations in the AI and the aMCC, which are the core regions of the empathy for pain network (Fan et al. 2011; Lamm et al. 2011). With regard to the effects of compassion training, we had the following hypotheses: Given that other-related feelings of warmth and concern are key components of compassion, we anticipated that loving kindness training would increase activity in the brain regions previously associated with love (Bartels and Zeki 2000; Aron et al. 2005) or affiliation (Vrticka et al. 2008; Strathearn et al. 2009), such as the (medial) orbitofrontal cortex (mOFC), the putamen, the middle insula, and the ventral tegmental area/substantia nigra (VTA/SN, which are mentioned together as the spatial resolution of the present fMRI data does not allow for an unambiguous distinctions between both midbrain structures). We furthermore expected these neural changes to be paralleled by an increase in positive affect and a decrease in negative affect.
Materials and Methods
Subjects
We investigated a total of 94 right-handed healthy female volunteers between 18 and 35 years of age (mean age = 24.3, standard deviation [SD] = 4.17) (Fig. 1A, see “Main Experiment, Participants” in Supplementary Material). The study was approved by the Research Ethics Committee of Zurich (“Kantonale Ethikkommission des Kantons Zürich—Spezialisierte Unterkommission Psychiatrie, Neurologie, Neurochirurgie”; E-25/2008) and was carried out in compliance with the Declaration of Helsinki. All subjects gave written informed consent, were paid for their participation, and were debriefed after the study was completed.
Measures
A new measure of social affect, the SoVT, was developed to assess social emotions in response to short videos of 10–18 s. To avoid habituation and recognition effects, participants were tested with a different randomly selected parallel video set at each measurement time (for details, see “Task Development” in Supplementary Material). This assured that all 3 video sets (A, B, and C) were equally viewed at all measurement time points by all groups of participants. The videos were presented without sound and depicted raw material cast for news or excerpts from documentaries. Each of the 3 video sets was composed of 12 HE videos which depicted persons in distress and 12 LE videos which served as a high-level baseline and showed people performing everyday activities (Fig. 1B). Videos were shown in blocks of 3 HE or 3 LE videos (resulting in a total of 24 videos per measurement point). Following each video, participants rated on 3 different scales how much empathy, positive affect, and negative affect they experienced when watching the videos (scale: from 0 to 10, presented in a pseudo-randomized order for 6 s each). Null events displaying a fixation cross were presented for 10 s after each mini-block. Participants were instructed to observe the videos and to pay attention to their emotional response. At post-training, participants were encouraged to make use of their trained competences when viewing the videos.
The memory task consisted of 3 parallel lists of 34 words, which enabled us to test participants with a different list at each measurement time point. Words were presented sequentially on a computer screen for 4 s each, followed by a 2-s fixation cross. Subsequently, subjects were given 5 min to fill in a table with the recollected words, if possible, in the correct sequence. To obtain questionnaire measures of affective traits, we measured empathy by means of the Interpersonal Reactivity Index (IRI, Davis 1983) and compassion by means of the Compassionate Love Scale (CLS, Sprecher and Fehr 2005).
Procedure
Pre-training measurements were collected 1–2 weeks prior to training and post-training measurements were collected 2–12 days after training. Both measurements consisted of behavioral and fMRI components and were essentially identical, apart from specific questionnaires collected only at pre- or post-training in the context of a related study (Leiberg et al. 2011). In the behavioral part of the study, participants played the Zurich Prosocial Game (Leiberg et al. 2011), performed the memory task, and completed a series of questionnaires. Prior to being scanned with the SoVT, participants received written and verbal instructions on the task and performed 3 practice trials. The total duration of the video task in the fMRI scanner was 15 min, consisting of two 7.5-min sessions separated by an anatomical scan, resulting in a total scan time of 20 min. To validate the SoVT, 33 participants were shown video set A, 31 viewed video set B, and 30 saw video set C at pre-training.
Compassion Training
The compassion training group attended a 1-day course of loving kindness meditation (6 h) led by a teacher with >10 years of teaching experience. The course was aimed at fostering benevolent and friendly attitudes toward oneself and other persons based on techniques from Eastern contemplative traditions (Salzberg 2002). Ultimately, the goal was to develop compassion as a generalized prosocial feeling and motivation, independent of particular persons or situations. The entire course was carried out in a quiet contemplative state adopted in an upright seated posture or during silent walking. Participants received a theoretical introduction, followed by guided practices (15–30 min) of strengthening feelings of warmth and care through the visualization of a close loved person (e.g., a child). Participants sequentially extended these feelings toward themselves, a close person, a neutral person, a person in difficulty, and finally toward strangers and human beings in general. At the end of the course, participants practiced modulating the intensity of compassion toward a loved person. After the day of training, participants filled out a debriefing questionnaire which asked several quantitative questions and gathered qualitative reports about the participants’ training experiences (see Supplementary Tables S1 and S2). These additional reports confirmed that, on average, participants liked the loving kindness training and could successfully engage in the different practices taught during that day. In between pre- and post-measurements, participants practiced the trained method during evening classes, during 3 laboratory measurements (45 min each) in the context of another study, and at home. Participants were asked to record their number of home training hours. Twenty-six of 28 participants in the compassion group returned their questionnaires to us indicating a mean of 5.7 practice hours (range 45 min to 11.17 h). We cannot be certain about the reliability of these data, as we did not use electronic measurement tools to objectively assess home practice. Furthermore, participants were encouraged to practice throughout their daily lives and, indeed, several reported engaging in the trained method in everyday life situations (e.g. using public transport). Finally, the number of training hours did not correlate with any of the dependent variables. Taken together, we know certainly that in addition to the one intense day of compassion training (6 h) and the informal training at home and in everyday life, participants practiced a minimum of 2 additional supervised training hours.
Memory Training
Like the compassion training group, the memory control group also attended a 1-day course (6 h) held by a teacher with >10 years of experience in teaching the method of loci, a technique to memorize items in an ordered sequence (Bower 1970). After initial practice with a self-chosen sequence of 10 locations, participants learned a fixed sequence of 34 locations (a route through Zurich). Lists of items were remembered by creating vivid mental images that associated each item with a location. Item lists were recalled by mentally revisiting each location on the route and by bringing to mind the previously created mental images. Thus, both training groups engaged in active mental imagery: The memory group trained a purely cognitive ability and the compassion group trained an emotion related to attachment and affiliation. The post-training debriefing questionnaire revealed that participants of the memory training group also enjoyed the mnemonic training and were also able to apply the taught memory technique (see Supplementary Table S1). As with the compassion group, the memory group also participated in evening classes or trained at home. Since the return rate of the self-reported training hours of the memory control group was not reliable (only 22 of 30 persons provided this information), these data were not analyzed.
Behavioral Data Analysis
Self-reports of empathy, positive affect, and negative affect in response to the SoVT, and performance in the memory task were analyzed using repeated-measures multivariate analyses of variance (MANOVAs; with Pillai's trace statistics), followed-up by analyses of variance (ANOVAs) and t-tests. All statistical tests were 2-tailed and used an alpha level of 0.05. Correlation coefficients were computed using Pearson's r. Correlations were compared using the Dunn and Clark's z test. At post-training measurement, in 4 participants the rating of 1 of 2 runs were lost due to a program error reducing subjective ratings from the initial 24 videos to 12 videos for these 4 participants. Since the recall data from the memory task were missing from 1 subject in the compassion group and from 1 subject in the memory group, the total sample size for the memory task was N = 56.
fMRI Data Acquisition and Analyses
General linear models and parametric modulations of ratings were estimated on the first level for each participant after standard preprocessing (realignment, normalization, and smoothing). The resulting contrasts were then entered into second-level random effects ANOVAs and t-tests (for details, see “Main Experiment, fMRI Data Acquisition and Analyses” in Supplementary Material). Neuroimaging results are reported at P < 0.05, corrected for family-wise errors (FWE) using Gaussian random fields theory and a cluster extent threshold of k = 5 voxels. When restricting the search volume to a priori defined ROIs, results are reported with a FWE-corrected significance threshold of P < 0.05 using small volume correction (SVC). For completeness, we also report brain regions activated at P < 0.001, uncorrected in the tables. Since these activations were not in the a priori defined ROIs and did not survive whole-brain correction, we do not draw conclusions about them in the present paper.
Results
Validation of the Socio-Affective Video Task
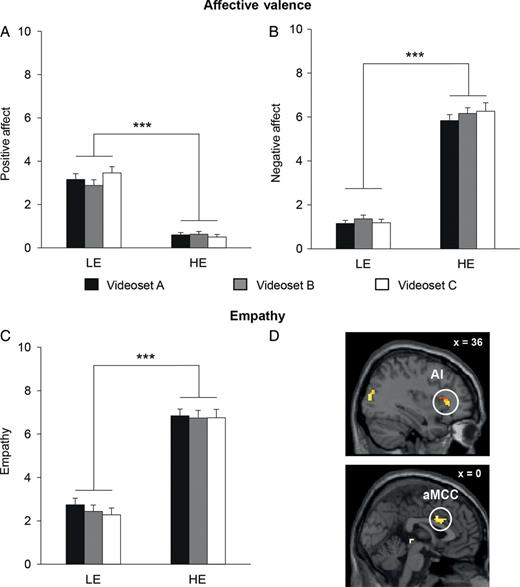
Analyses focusing on the validity of the SoVT confirmed the following (for details, see “Task Development” in Supplementary Material, Supplementary Fig. S2, and Supplementary Tables S3–S5): Watching HE as compared with LE videos evoked more negative and less positive feelings. Furthermore, participants reported feeling more empathy toward other people in distressing situations compared with everyday situations (Fig. 2A–C). Pearson correlations between self-report ratings on the SoVT and questionnaire measures of empathy (assessed by the IRI, Davis 1983) and compassion (measured by the CLS, Sprecher and Fehr 2005) confirmed the external validity of the SoVT. Empathy ratings for HE videos were significantly correlated with total empathy scores (P < 0.05, r = 0.23). This effect was driven by the subscales empathic concern (P < 0.01, r = 0.34) and fantasy (P < 0.001, r = 0.42). Compassionate love was correlated with empathy for LE and HE videos (P < 0.01, r = 0.29 and P < 0.001, r = 0.36, respectively), as well as with positive affect for LE (P < 0.01, r = 0.29) and HE videos (P < 0.05, r = 0.26).
{kind=link}
Subjective emotional experiences and related neural activations evoked by the SoVT. (A–C) Average subjective ratings of positive affect, negative affect, and empathy in response to LE and HE video stimuli at pre-training (zero, no affect; 10, maximum affect). Whereas the 3 video sets (A–C) did not differ on any of the ratings, the effect of video type (LE and HE videos) was highly significant for each dependent variable. Error bars indicate the standard error of mean (SEM); ***P< 0.001. (D) SVC with a functional activation map from a recent meta-analysis on empathy for pain (Lamm et al. 2011) revealed that activations in the AI (x = 34, y = 26, z = 2; Z = 4.14, P < 0.05, SVC) and the aMCC (x = 0, y = 10, z = 34; Z = 4.66, P < 0.05, SVC) were parametrically modulated by subjectively experienced empathy for HE videos during the SoVT. Color-coded activations with brighter colors indicating lower P values were rendered on a Montreal Neurological Institute (MNI) template in the neurological orientation. Inset x values indicate a stereotactic coordinate of the shown slice in the MNI space. AI, anterior insula and aMCC, anterior medial cingulate cortex.
Neuroimaging results (reported at P < 0.05, FWE-corrected) revealed that HE videos compared with LE videos, as well as parametric modulations of negative affect and empathy, induced activations in a network comprising areas in the occipital, temporal, and parietal lobes, as well as in the subcortical areas including the amygdala, thereby replicating previous findings on neural activity typically involved in the processing of negative pictures or videos (Goldin et al. 2008; Vrticka et al. 2011). Conversely, LE compared with HE videos elicited stronger activation in the posterior insula, the mOFC, the fusiform, and the precentral gyrus. In contrast to negative affect ratings, positive affect ratings mapped to the mOFC, a brain region consistently implicated in the experience of hedonic feelings as well as in the encoding of value and reward (Kringelbach and Berridge 2009; Rangel and Hare 2010; Grabenhorst and Rolls 2011). Finally, the validity of the SoVT as a measure of empathy for suffering in others could be confirmed by showing that neural activity parametrically modulated by empathy ratings in response to HE videos mapped to activations in the AI (x = 34, y = 26, z = 2; Z = 4.14, P < 0.05, SVC), and the aMCC (x = 0, y = 10, z = 34; Z = 4.66, P < 0.05, SVC)—both core regions involved in empathy for others’ suffering (Fan et al. 2011; Lamm et al. 2011, Fig. 2D and Table S5).
Training-induced Plasticity
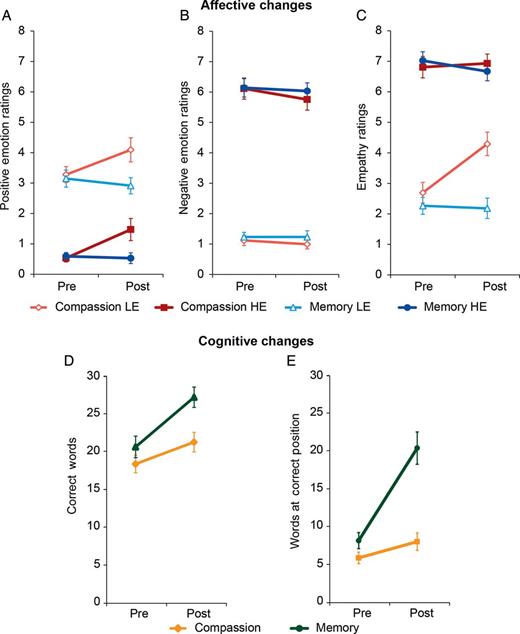
To assess whether short-term compassion training would lead to increased positive and decreased negative affect during vicarious experiences, we performed a 2 × 2 × 2 repeated-measures MANOVA with video type (HE and LE videos) and time (pre- and post-training) as within-subject factors and training group (compassion and memory) as a between-subject factor. The dependent variables were empathy, positive affect, and negative affect ratings. The main effects of video type (F3,54 = 320.70, P < 0.001, η² = 0.95) and time (F3,54 = 2.80, P < 0.05, η² = 0.14) were significant, and the main effect of training group showed a trend toward significance (F3,54 = 2.70, P = 0.06, η² = 0.13). Significant interactions were found for video type × time (F3,54 = 4.70, P < 0.01, η² = 0.21), video type × group (F3,54 = 3.27, P < 0.05, η² = 0.15), and time × group (F3,54 = 5.64, P < 0.01, η² = 0.24); the triple interaction video type × time × group was not significant (F3,54 = 2.11, P = 0.11). As expected, the difference between HE and LE videos was confirmed for all 3 dependent variables in paired t-tests (all t57 ≥ 15.18, all P < 0.001). Univariate ANOVAs showed that the main effect of time was significant for empathy (F1,56 = 4.57, P < 0.05) and positive affect (F1,56 = 4.36, P < 0.05). Furthermore, all 3 interactions were significant for the dependent variable empathy (all F1,56 ≥ 8.15, all P < 0.01). Positive affect only showed a time × group interaction (F1,56 = 8.66, P < 0.01), whereas contrary to our expectations, no training effects were found for negative affect. Follow-up paired t-tests revealed that in the compassion group, empathy ratings for LE videos were higher at post- than at pre-training (t27= 3.71, P < 0.01) and independent t-tests showed that post-training empathy ratings for LE videos were higher in the compassion than in the memory group (t56= 4.18, P < 0.001). Similarly, positive affect ratings were higher at post- than at pre-training in the compassion group (t27= 2.68, P < 0.05) and independent t-tests determined that positive affect ratings at post-training were higher in the compassion than in the memory group (t42.26= 2.72, P < 0.05). All other effects were not significant. In summary, compassion training (but not memory training) led to significant increases in ratings of positive affect for LE and HE videos and to an increase in empathy ratings for LE videos (Fig. 3A–C). Of note, self-reports of 26 of 28 participants from the compassion group in a post-scan debriefing indicated that when viewing the videos after the training, participants seemed to have adopted a compassionate stance toward HE videos (mean = 5.38, SD = 0.88) and LE videos (mean = 4.26; SD = 1.38, on a scale from 1, minimum to 7, maximum). This suggests that these participants indeed engaged in compassion when viewing the SoVT at post-training.
{kind=link}
Behavioral effects of compassion and memory training. (A) Self-reported positive affect in response to LE and HE videos increased after compassion training, but not after memory training. (B) No significant changes were observed for negative affect. (C) Compassion, but not memory training, increased empathy toward people in LE videos. (D) Memory and compassion training improved the number of correctly remembered words. Importantly, at post-training measurement, the memory group remembered significantly more correct words than the compassion group. (E) Memory training induced an increase in the number of words remembered in the correct position. Error bars indicate the SEM.
Interestingly, in the compassion group the correlation between empathy and negative affect at pre-training measurement was reduced at post-training, which may be accounted for by the increase in empathy ratings in response to LE videos (pre-training: r = 0.69, P < 0.001; post-training: r= 0.15, P = 0.44; Dunn and Clark's z test Z = 2.96, P < 0.01). No such effect was observed for the memory group (pre-training: r = 0.65, P < 0.001; post-training: r = 0.82, P < 0.001; Z = 1.04, P = 0.3), and no other correlations showed significant changes. Furthermore, the effectiveness of the memory control training was confirmed (see “Analysis of the Memory Task” in Supplementary Material and Fig. 3D,E).
As the field of compassion research is just emerging, we performed multiple experiments to assure replicability, validity, and generalizability of the training based neuroimaging results. Prior to conducting the main experiment, we performed 3 studies to determine the core neural signatures underlying the generation of compassionate feelings. In particular, these independent experiments included different populations (novice meditators and an expert practitioner) across different countries (the Netherlands and Switzerland) using different task parameters (immersion in compassionate states without an external task and brain responses to the SoVT). Through such a multi-study and multi-method approach, we aimed to reduce the measurement noise, secure replicability and validity, and optimize the generalizability of the findings. First, to obtain external validity, we tested a long-term expert practitioner (>35 years of daily practice in compassion-enhancing mental training techniques), while he immersed himself in 3 different compassionate states during 15 fMRI sessions acquired over 3 days (see “Expert Data” in Supplementary Material). We observed that across all 3 compassion-enhancing techniques, the generation of strong, when compared with weak, feelings of compassion consistently increased activations in subcortical structures including the caudate nucleus and striatum, and cortical regions, including the mOFC and supplementary motor areas (P < 0.05, FWE-corrected; Supplementary Table S6). In a next step, we derived independent ROIs based on 2 previously conducted short-term compassion training experiments in meditation-naive participants. Specifically, the first experiment focused on the neural correlates reflecting subjective intensity ratings of newly trained compassionate states, while the other experiment measured neural changes related to compassion training in responses to others’ suffering in the newly developed SoVT (total N = 46, see “ROI construction” in Supplementary Material and Supplementary Table S7). Interestingly, the neural effects of short-term compassion training were consistent with activations in the expert practitioner in the right mOFC, VTA/SN, pallidum, and putamen (Fig. 4 orange boxes), as well as bilaterally in the precentral gyrus, amygdala, and thalamus and on the left side in the pallidum, putamen, middle occipital gyrus, supramarginal, and superior temporal gyrus.
![Effects of compassion and memory training on neural responses to HE videos. The contrast [Compassion ΔHE > Memory ΔHE] revealed activations (P < 0.05; SVC) in (A) the right mOFC, (B) the right VTA/SN, (C) the right pallidum, and (D) the right putamen. Bar charts show the change in parameter estimates in the depicted independent region of interest; error bars denote the SEM. Orange boxes show neural activations (P < 0.05; FWE-corrected) of an expert practitioner immersed in 3 compassionate states [high > low degree] across 15 sessions depicted within independent ROIs. Color-coded activations with brighter colors indicating lower P-values were rendered on an MNI template in neurological orientation. Inset x/z values indicate a stereotactic coordinate of the shown slice in the MNI space.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/cercor/23/7/10.1093_cercor_bhs142/2/m_bhs14204.jpeg?Expires=1716358202&Signature=1-EzRTAGKaHCY3JCfuD7ICcq71Qyd6tQE8TkdJoVxNAvJcnB0wzBoWV0q2XannMYM2FfTjdLwilmkBmbbl7fw6D327dNd~mg5CZqxaVVfMU6VkCZ96wGcCOwZVQ-62caVJpxR5jSlnE~xam58MWdif71IJeq4XKPNkgz52unD~yIU89EX90SrZ8YTq0plrct0LmXwH6T6KL2mt7y8AkiDeJKR~FYPvQcdfpX4hjLhaM6~-GJM1GilJJob-7DCw~UjTBmnm4-H1IYsHLBsCoWPmBGO8u~1RfQlcrNuCUYmLb5SskK7mrrYEYwwqInEaTfQfKUnbDJLzyOUzLYAV7e-w__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
{kind=link}
Effects of compassion and memory training on neural responses to HE videos. The contrast [Compassion ΔHE > Memory ΔHE] revealed activations (P < 0.05; SVC) in (A) the right mOFC, (B) the right VTA/SN, (C) the right pallidum, and (D) the right putamen. Bar charts show the change in parameter estimates in the depicted independent region of interest; error bars denote the SEM. Orange boxes show neural activations (P < 0.05; FWE-corrected) of an expert practitioner immersed in 3 compassionate states [high > low degree] across 15 sessions depicted within independent ROIs. Color-coded activations with brighter colors indicating lower P-values were rendered on an MNI template in neurological orientation. Inset x/z values indicate a stereotactic coordinate of the shown slice in the MNI space.
Finally, we analyzed functional neural plasticity induced by compassion training compared with memory training within the independently derived ROIs (Supplementary Table S8). To this end, we computed first-level contrast images of post- > pre-training activation separately for LE and HE videos (ΔLE and ΔHE videos) and entered these into a 2 × 2 repeated-measures random effects ANOVA with the within-subject factor video type (ΔLE and ΔHE videos) and the between-subject factor group (compassion and memory training). We first tested the effect of the group across both ΔLE and ΔHE videos. This revealed that the compassion group had stronger activations in the right putamen (x = 28, y = −6, z = 6; Z = 3.08, P < 0.05, SVC) and pallidum (x = 26, y = -8, z = 4; Z = 2.88, P < 0.05, SVC). The interaction [(Compassion ΔHE − ΔLE) > (Memory ΔHE − ΔLE)] revealed activations in the right mOFC (x = 12, y = 38, z = -18; Z = 2.73, P < 0.05, SVC). As the main focus of this study was to determine whether compassion training was able to alter brain responses elicited when exposed to the distress of other people, we also directly contrasted training-related changes when watching HE videos between the compassion and the memory group. This revealed that compared with memory training, compassion training was associated with activations in the mOFC, VTA/SN, putamen, and pallidum (all right lateralized, P < 0.05, SVC; Fig. 4). Furthermore, training-related changes were not significant in response to LE videos, although Figure 4 suggests that there was a trend in the same direction as for HE videos.
Interestingly, the memory group showed increased neural activity in response to LE videos in the right hippocampus (x = 40, y = −16, z = −22; Z = 2.57, P < 0.05, SVC), a region relevant for memory processes (Squire et al. 2004) and the right fusiform gyrus (x = 40, y = −16, z = −24; Z = 2.71, P < 0.05, SVC). No other training-based activation changes survived correction in the compassion-related ROIs.
Parametric analyses on the changes in ratings revealed that the overall increase in positive ratings (across HE and LE videos) in the compassion group correlated with activity changes in the right putamen (x = 34, y = −2, z = −2; Z = 2.99, P < 0.05, SVC). Changes in negative affect ratings correlated with activations in the left supramarginal gyrus (x = −50, y = −26, z = 20; Z = 2.8, P < 0.05, SVC), the left superior and the left middle temporal cortices (x = −44, y = −44, z = 10; Z = 2.99 and x = −46, y = −46, z = 8; Z = 3.14, respectively, both P < 0.05, SVC), while no correlations were observed with changes in empathy ratings.
Discussion
The present study investigated functional neural plasticity induced by compassion training. In particular, we studied which subjective and neural changes in response to distressed others were induced by compassion or memory training. As fMRI compatible paradigms for the repeated probing of affective responses to social situations are rare, we developed and validated a new fMRI task, the SoVT. The SoVT was optimized for repeated measurements: We generated 3 parallel video sets which could be used interchangeably. The employed videos depicted short everyday life scenes of people in low or high emotional states. Self-reports collected after each video allowed the dissociation of empathy, positive and negative affect in response to other people's suffering.
Essentially, we observed that compassion training compared with memory training: 1) increased positive affect, even in response to others’ suffering, and 2) was associated with stronger activations in a specific neural network including the mOFC, the pallidum, the putamen, and the VTA/SN—brain regions previously implicated in positive valuation (Kringelbach and Berridge 2009), as well as love (Bartels and Zeki 2000, 2004; Aron et al. 2005; Beauregard et al. 2009) and affiliation (Vrticka et al. 2008; Strathearn et al. 2009). These results are particularly striking, since participants were exposed to aversive (HE) videos, which at pre-training actually elicited negative affect and empathy-related activations in the AI and the aMCC—core regions of the empathy for pain network (Fan et al. 2011; Lamm et al. 2011) that have typically been modulated by subjective experience of unpleasantness (Rainville 2002). Finally, the parallel nature of the neural pattern in several experiments including novices and an expert, after lends further support for the replicability and validity of the observed compassion-related brain activation pattern.
In light of previously reported findings, the observed activation pattern underlying the training of compassion accords with neural correlates of love, affiliation, and positive affect. More precisely, romantic (Bartels and Zeki 2000; Aron et al. 2005) and maternal love (Bartels and Zeki 2004) have been shown to induce stronger activations in the VTA/SN, the middle insula, the putamen, and the pallidum. Similarly, activations in the striatum and the VTA were observed in response to smiling faces and this activation was reduced in individuals with avoidant attachment (Vrticka et al. 2008). In addition, secure attachment in mothers seeing photographs of their happy children has been related to activity in the ventral striatum and the mOFC (Strathearn et al. 2009). More generally, mOFC activations were consistently associated with positive affect, such as reward value and pleasure (Kringelbach and Berridge 2009; Rangel and Hare 2010; Grabenhorst and Rolls 2011), which ties in well with the pre-training observation of a correlation between ratings of positive affect and mOFC activity. Furthermore, the present results are in line with cross-sectional studies focusing on compassion either through instruction-induced effects in naive participants (Beauregard et al. 2009; Kim et al. 2009) or through testing expert meditation practitioners (Lutz et al. 2008). For example, the involvement of VTA/SN and mOFC was also observed when participants were instructed to adopt a compassionate attitude toward sad faces (Kim et al. 2009) and activations in the VTA, the pallidum, and the middle insula were observed when adopting an attitude of love toward disabled persons (Beauregard et al. 2009). Middle insula activation was reported to be crucial when expert practitioners immersed themselves in compassion meditation while hearing human cries of distress (Lutz et al. 2008). Note that in the present study, middle insula activation was also observed to be parametrically modulated by changes in positive affect ratings in the compassion group, albeit at a lower threshold of P = 0.001. The present results extend the previous cross-sectional findings in an important way as they provide, for the first time, longitudinal evidence that activations in the mOFC, the VTA/SN, the pallidum, and the putamen (extending into the middle insula) are stronger in response to distressing video stimuli after compassion training, but not following memory training.
Neural changes were particularly pronounced for HE videos, although the parameter estimates in Figure 4 show similar but weaker changes in response to LE videos. On the behavioral level, this effect was paralleled by increased self-report ratings of positive affect in response to both HE and LE videos. The overall increase in positive affect through the cultivation of warmth and benevolence in the present study adds to the finding that daily self-reports of positive emotions were augmented after loving kindness training (Fredrickson et al. 2008). Extending previous results, the present study actually shows that positive emotions were not only increased in response to everyday life situations, but even in response to witnessing the distress of others. The observed increase in reported positive affect—even when exposed to the suffering of others—suggests that persons trained in compassion can encounter social situations in general and distressing situations in particular with positive, other-oriented affect. A review of the qualitative self-report data collected after the training day of the compassion group gave us some more information about the qualia associated with the generation of loving kindness and compassion (see Supplementary Table S2). For instance, participants indicated that they liked, “the feeling of warmth”; “the wonderful, fulfilling feeling to wish others well”; “feelings of love, security, shelter”; “to think of other people and […] give them kindness”; “a feeling of happiness that arises.” They further reported, “I felt an enormous heat. It was nice to feel that one could create something like that inside oneself.”; “It was easier than I thought to generate feelings of benevolence toward a stranger.” Although these qualitative self-reports cannot be quantified and used in analyses, they nonetheless give us valuable indications that loving kindness training strengthened feelings of warmth and care toward oneself and others. Finally, the indication that participants indeed used the practice of loving kindness when exposed to the SoVT at post-training measurement, suggest that participants re-interpret the meaning of empathy: instead of experiencing empathy as a distressing and aversive emotion that may ultimately result in withdrawal tendencies (Batson et al. 1983; Eisenberg et al. 1989), compassion training may have induced an understanding of empathy as a rather positive feeling associated to empathic concern (Batson 2009) and warmth.
Finally, contrary to our expectation, compassion training only increased positive affect, but did not decrease negative affect. In line with previous reports of co-activated positive and negative affects (e.g., Larsen et al. 2001), our finding underlines the need to measure positive and negative affects independently (see also Watson et al. 1988; Cacioppo and Berntson 1994). Indeed, the co-activation of positive and negative affect may be a crucial prerequisite for successful coping with stressors (Larsen et al. 2003). Future studies may help to dissociate the neural substrates underlying positive and negative affective experiences associated with social and vicarious emotions.
In a broader context, the current results extend our understanding of existing strategies known to be useful for coping with negative affect such as extinction learning (Gottfried and Dolan 2004) or different forms of explicit emotion-regulation strategies such as cognitive reappraisal (Ochsner et al. 2002; Kalisch 2009), emotion suppression (Levesque et al. 2003; Phan et al. 2005), or detachment (Walter et al. 2009). In line with previous reports on beneficial effects of compassion training on mood (Fredrickson et al. 2008) and health (Pace et al. 2009), the current data show that responding to potentially distressing situations with compassion benefits the person who experiences it through strengthening positive affect. Furthermore, compassion benefits others through fostering prosocial behavior (Leiberg et al. 2011). By consequence the generation of compassion may not only help others, but also be beneficial for oneself by providing a potentially powerful coping strategy. However, to situate compassion in relation to the existing framework of emotion regulation strategies, future studies are needed which directly compare different emotion regulation techniques with compassion generation.
Regarding the duration of training, the current study ranged at the lower end of interventions, therefore examining short-term functional neural plasticity. Future studies are needed to map out the behavioral, subjective, and neuronal changes that occur after longer compassion training interventions of several months to years. In addition, it would be interesting to understand the role specific neurotransmitter systems play in strengthening compassion. Although the spatial resolution used in the present experiments did not allow us to disentangle the VTA and the SN, the relatively stronger activation for compassion compared with memory training in this area is interesting, since the VTA and the SN are rich in dopamine (Fields et al. 2007) and the SN is rich in oxytocin binding sites (Loup et al. 1989). In addition, the VTA is a key region mediating the rewarding effects of opiates (Burgdorf and Panksepp 2006). Future neuropharmacological intervention research and Positron Emission Tomography with specific ligands might help to unravel the contribution and interaction of dopaminergic reward (Schultz 2010), oxytocin-related affiliative processes (Strathearn et al. 2009), and opiate-mediated well-being (Burgdorf and Panksepp 2006) in the generation of compassion.
In conclusion, the present findings provide evidence for functional neural plasticity induced by compassion training and thus constitute a crucial step in the endeavor of understanding neural plasticity in the affective domain. The results suggest that compassion training can serve as a new, powerful method for enhancing positive affect in response to adverse situations. This is in accordance with findings of beneficial effects of compassion training for strengthening personal resources (Fredrickson et al. 2008) and health (Pace et al. 2009). As positive affect has been found to be associated with resilience (Fredrickson et al. 2003), future studies should explicitly address whether compassion training can serve as a buffer against stress, depression, and anxiety, and which physiological substrates mediate this effect. Importantly, in comparison to other techniques for increasing positive affect, compassion benefits both, the person who experiences it (through strengthening positive affect) and the recipient of compassion (through fostering prosocial motivation). In the long-run, this promises to have major implications for the development of interventions for normal and clinical populations.
Funding
This work was supported by the University of Zurich (“Neural Foundations of Empathy Learning”, Forschungskredit, grant number 53990601 to O.M.K.), grants to T.S. from the Neuroscience Center Zurich, the Swiss National Science Foundation (SNF NCCR Affective Sciences: “Neuronal and developmental mechanisms of human empathy”; SNF NCCR Neural Plasticity and Repair: ”Plasticity of the empathic and attentional brain”), the Foundation for Research in Science and Humanities at the University of Zurich (“Neural foundations of empathy training”), and a Francisco J. Varela Research Award to S.L. from the Mind and Life Institute. The research leading to these results has also received funding from the European Research Council under the European Community's Seventh Framework Program (FP7/2007-2013/ ERC Grant agreement number 205557 to T.S.).
Notes
We are deeply grateful to Dr Matthieu Ricard for all the fruitful discussions that resulted in this research. Furthermore, we gratefully acknowledge Dr Rainer Goebel and Dr Bettina Sorger for their contribution in Maastricht. Finally, our gratitude goes to the teachers Catherine Felder, Annette Rentz-Lühning, Jotika Hermsen, and Ariya Ñani, as well as to Fred von Allmen. Conflict of interest: none declared.