





Abstract
Urbanization increasingly exposes birds to multiple sources of direct anthropogenic mortality. Collisions with buildings, and windows in particular, are a top bird mortality source, annually causing 365–988 million fatalities in the United States. Correlates of window collision rates have been studied at the scale of entire buildings and in relation to the surrounding landscape, and most studies have only assessed correlates for all birds combined without considering season- and species-specific risk factors. In Stillwater, Oklahoma, USA, we conducted bird collision surveys at 16 buildings to assess building structural-, vegetation-, and land cover-related collision correlates. Unlike past studies, we focused at the scale of individual building façades, and in addition to considering correlates for total collisions, we assessed correlates for different seasons and separately for 8 collision-prone species. Several façade-related features, including proportional glass coverage, façade length, and façade height, were positively associated with total collisions and collisions for most separate seasons and species. Total collisions were also greater at alcove-shaped façades than flat, curved, and portico-shaped façades. We found that collision correlates varied among seasons (e.g., surrounding lawn cover important in summer and fall, but not spring) and among species (e.g., surrounding impervious cover positively and negatively related to collisions of Painted Bunting [Passerina ciris] and American Robin [Turdus migratorius], respectively). Given the importance of glass proportion, collision reduction efforts should continue to focus on minimizing and/or treating glass surfaces on new and existing buildings. Our species- and season-specific assessments indicate that management of some collision risk factors may not be equally effective for all seasons and species. Future research, policy, and management that integrates information about collision risk for all bird species and seasons, and at multiple scales from building façades to the surrounding landscape, will be most effective at reducing total mortality from bird–window collisions.
Resumen
La urbanización expone cada vez más a las aves a múltiples fuentes de mortalidad antropogénica directa. Las colisiones con edificios, y con ventanas en particular, son una de las principales fuentes de mortalidad de aves, causando anualmente 365–988 millones de fatalidades en Estados Unidos. Las correlaciones de las tasas de colisión contra ventanas han sido estudiadas a escala de edificios enteros y en relación con el paisaje circundante, y la mayoría de los estudios solo han evaluado las correlaciones para todas las aves en conjunto, sin considerar factores de riesgo vinculados específicamente con la estación y la identidad específica. En Stillwater, Oklahoma, EEUU, realizamos muestreos de colisiones de aves en 16 edificios para evaluar las correlaciones de las colisiones relacionadas con la estructura del edificio, la vegetación y la cobertura del suelo. A diferencia de estudios anteriores, nos centramos a la escala de fachada de edificios individuales, y además de considerar las correlaciones para colisiones totales, evaluamos las correlaciones para diferentes estaciones y de modo separado para ocho especies con tendencia a colisionar. Varios rasgos relacionados con las fachadas, incluyendo la proporción de cobertura de vidrio y el ancho y la altura de la fachada, estuvieron positivamente asociados con las colisiones totales y con las colisiones para la mayoría de las estaciones y de las especies por separado. Las colisiones totales también fueron mayores en las fachadas en forma de alcoba que en las fachadas planas, curvas o en forma de pórtico. Encontramos que las correlaciones con las colisiones variaron entre estaciones (e.g., la cobertura de pasto circundante fue importante en verano y otoño, pero no en primavera) y entre especies (e.g., la cobertura impermeable circundante se relacionó positiva y negativamente a las colisiones de Passerina ciris y Turdus migratorius, respectivamente). Dada la importancia de la proporción de vidrio, los esfuerzos destinados a la reducción de las colisiones deberían continuar enfocándose en minimizar y/o en tratar las superficies de vidrio de los edificios nuevos y existentes. Nuestra evaluación específica por especie y estación indica que el manejo de algunos factores de riesgo de colisión puede no ser igualmente efectivo para todas las estaciones y especies. Las investigaciones, políticas y manejos futuros que integren información sobre el riesgo de colisión para todas las especies de aves y estaciones, y a múltiples escalas desde la fachada del edificio hasta el paisaje circundante, serán más efectivas para reducir la mortalidad total originada por las colisiones de las aves contra las ventanas.
Introduction
Urbanization is increasing rapidly with urban land cover expected to triple globally from 2000 to 2030 (Seto et al. 2012). Urbanization restructures biotic communities because tolerance to urban development is variable among species, and urban landscapes are variable with regard to abiotic conditions (e.g., temperature), vegetation cover, and human-built features (Faeth et al. 2005, Fischer et al. 2015, Oliveira Hagen et al. 2017). Even urban-avoiding species sometimes traverse urban landscapes during migration or other major movements, which causes many species to interact at least briefly with urban environments (Pennington et al. 2008, Seewagen et al. 2010, Homayoun and Blair 2016). Birds in urban settings, including migratory species that otherwise spend little time in heavily developed areas, are vulnerable to building collision mortality, a major conservation issue that has increasingly been the focus of scientific, management, policy, and public attention (Avery 1979, Erickson et al. 2005, Klem 2015, Seewagen and Sheppard 2017). Such collisions largely occur at windows and cause an estimated 365–988 million bird fatalities annually in the United States (Loss et al. 2014).
Bird responses to the spatial heterogeneity of resources like food and cover, and the built environment, occur at multiple scales and influence spatial variation in movements, habitat use, and thus bird–window collision risk (Hager et al. 2017). At broad scales, factors like proximity to water and extent of urbanization may affect attraction of birds to the general area surrounding a building. Artificial night lighting associated with urban areas can also be a broad-scale attractant that draws nocturnally migrating birds into urban areas (Horton et al. 2019) and nearer to human-built structures including buildings (Van Doren et al. 2017, Winger et al. 2019). At fine scales, features of buildings (e.g., building height, adjacent vegetation) and individual building façades (e.g., façade shape, proportion covered by windows) likely influence collision risk for birds already present near buildings. Previous studies of bird–window collisions have focused on collision correlates operating at the scale of entire buildings or the broader landscape (O’Connell 2001, Hager et al. 2008, 2013, 2017; Gelb and Delacretaz 2009, Klem et al. 2009, Borden et al. 2010, Bayne et al. 2012, Cusa et al. 2015, Ocampo-Peñuela et al. 2016), even though limited descriptive research indicates collision risk can vary among façades within a single building (Cusa et al. 2015, Kahle et al. 2016). The few studies that have considered effects of façade-level characteristics (Klem et al. 2004, Borden et al. 2010, Bracey 2011, Cusa et al. 2015) have focused on a single factor (e.g., façade aspect) or used the term façade ambiguously, where it was unclear if the term described a specific section or the entire exterior surface of a building. Furthermore, even in studies that spanned multiple seasons, season-specific assessments of collision risk factors have rarely been conducted (but see Loss et al. 2019).
Regardless of scale, important correlates of window collision risk likely vary among bird species—because species differentially use resources, select habitat, and respond to the urban built environment—which likely contributes to the known variation in window collision risk in association with phylogeny and life history traits (Loss et al. 2014, Sabo et al. 2016, Wittig et al. 2017, Nichols et al. 2018). A study in Toronto, Canada, found that feeding guild and habitat preference of a species affected landscape-level correlates of window collision risk; collisions for foliage-gleaning species breeding in forested areas were positively correlated with the amount of greenspace around buildings while collisions for open woodland–inhabiting, ground-foraging species were positively correlated with urbanization (Cusa et al. 2015). A study in downtown Minneapolis, Minnesota, USA, assessed correlates of collisions for 5 different bird species (Loss et al. 2019); although glass area and/or amount of vegetation near buildings positively influenced collisions for all 5 species, there was some among-species variation in collision correlates. For example, collisions of Common Yellowthroat (Geothlypis trichas) were positively related to the amount of vegetation within 100 m of buildings while collisions for the other 4 species were related to vegetation within 50 m. Other than these examples from 2 major cities, assessments of species-specific correlates of window collisions are lacking. Thus, in addition to the need for formal research into building façade–level collision correlates, there is also a need to investigate species-specific correlates of collision risk.
Façade-scale and season- and species-specific assessments would be useful for informing management efforts to reduce bird–window collisions, such as considering collision risk in pre-construction building designs and mitigating collisions at existing buildings (e.g., by adding screens, cords, UV tape or paint, or patterned adhesive films and markers; Klem and Saenger 2013, Rössler et al. 2015, Menacho-Odio 2018, Sheppard 2019). Because mitigation across an entire building or multiple buildings may be cost-prohibitive, and in some cases unwarranted given evidence that collisions do not occur uniformly across all building façades, fine-scale understanding of collision correlates could help target management efforts in which only specific portions of buildings are treated. Given the lack of formal analyses regarding fine-scale correlates of bird–window collisions, our objective was to assess how bird–window collision rates in a small urban area in the central United States are influenced by façade-level variables, both within and across seasons and bird species. We addressed this objective by conducting near-daily surveys of 16 buildings in spring, summer, and fall to document specific collision locations and relating façade-specific collision rates to 8 potential predictor variables similar to factors known to influence collisions at coarser scales (e.g., building size, amount of glass, and nearby vegetation). Generally, we expected to observe more collisions at building façades that were larger, covered by a greater proportion of glass, closer to trees, and surrounded by more non-lawn vegetation cover. However, given differences in species life histories (e.g., migration strategy and timing) and other factors that vary seasonally (e.g., vegetation), we expected to find variation in collision correlates among species and seasons.
METHODS
Study Area and Study Design
We surveyed for bird carcasses at 16 buildings in Stillwater, Oklahoma, USA, a small urban area with a human population of ~46,000 and with the vast majority of buildings consisting of residences or small (<5 stories in height) office-type structures. Stillwater is in the Cross Timbers transitional ecoregion of the south-central United States, where deciduous forests from the east mingle with grasslands from the west to create a mixture of prairie, savannah, and woodland. We used a stratified approach to select survey buildings based on building size and amount of surrounding vegetation (Hager et al. 2017), but building selection was not completely random as we were constrained by building accessibility. The surveyed buildings included detached residences (n = 2), commercial off-campus-structures (n = 3), and classroom, office, and athletics buildings (n = 11) on the Oklahoma State University (OSU) main campus. We surveyed ≥6 days per week at all buildings from April 1 to October 31 in 2015 and in 2016 and at 14 buildings (excluding the 2 residences) from April 1 to May 31 in 2017. For seasonal delineations, we considered spring to be April to May, summer to be June to August, and fall to be September to October.
Collision Locations
Each survey consisted of a single observer walking slowly along the exterior perimeter of a single focal building, intensively searching a 2-m swath along all windowed walls. We also entered 3 buildings to survey ledges below windows that could not be observed from the outside. We alternated the direction that a building perimeter was surveyed on a daily basis (clockwise on even days, counter-clockwise on odd days) to minimize detection effects related to the angle and direction from which an observer could approach a carcass (e.g., obstacles, shading). Upon discovery of an intact carcass, we took photographs and recorded the location and a description of the carcass. We similarly documented remains indicative of a carcass, which usually consisted solely of feathers (i.e., feather pile) that had been plucked from the carcass by a scavenger. To avoid counting adventitiously lost feathers, we only counted feather piles that consisted of ≥5 feathers within a circular area ~15 cm in diameter.
To avoid counting a single carcass more than once, we collected bird remains using a plastic, sealable bag, and subsequently stored them in a freezer with unique alphanumeric identification codes. When we could not collect carcasses because they were on inaccessible ledges above ground level, and at one building where carcasses were regularly left as found as part of a concurrent study, we tracked the carcass condition and location to avoid double counting it on future surveys.
We used written observer descriptions to record locations for carcasses and non-fatal collisions in Google Earth Pro 7.3.2 (Google, Mountain View, California, USA), which allowed location accuracy to within ~2 m. When written location descriptions were ambiguous, we followed up within 1 day to have the observer who detected the carcass clarify the precise location by marking it on a map. We imported these collision locations to ArcGIS 10.2.1 (ESRI, Redlands, California, USA) to generate and analyze spatial data. To generate façade-specific carcass counts, we spatially joined carcass locations, both including and excluding feather piles, to polygons representing the 2-m-wide search area for each façade. We repeated this for each season (spring, summer, and fall) and for each species with ≥15 collision observations.
Façade Variables
We defined a façade as a distinct section of a building, typically bounded by corners, that was qualitatively homogenous and exhibited minimal variation in measurable traits like height. We characterized 8 façade-level variables, including façade height (m), façade length (m), façade type, distance to trees (m), 3 land cover variables, and proportion of the façade consisting of glass. We used digital photographs taken with a Panasonic DMC-ZS1 camera (focal length: 4.1–49.2 mm) and analyzed in ImageJ 1.48 (Schneider et al. 2012) to measure the height, length, area, and glass-covered area of each surveyed building façade (glass proportion was calculated as glass-covered area divided by total façade area). We took photographs perpendicular to the center of each façade at a height of ~2 m and from as far away as possible (range: 5–75 m) while still capturing the façade with minimal obstacles. To serve as a known-dimension reference for calibrating measurements of façades and windows in photographs, we directly measured ≥1 reference object (e.g., width of single window pane) at ~2 m height that was (1) clearly visible in the photograph, (2) near the center of the façade, and (3) occurred at multiple heights or along the length of the façade. Tall, long, or curved façades became non-orthogonal near the edges in photographs of the entire façade. To avoid biased measurements using these distorted portions of images, we used the known-length reference objects to adjust measurements incrementally away from the façade center. For very long façades, we took 2 photographs, each perpendicular to the façade at locations approximately one-fourth and three-fourths of the way along the façade’s total length, and we then combined area estimates generated for each half of the façade.
Façade type included 5 categories (Figure 1): (1) alcove, where the façade, usually in conjunction with the adjoining façades, formed an indentation/concavity in the building outline when viewed from above; (2) portico, where the façade formed an indentation/concavity in the building surface along a vertical plane resulting in a covered walkway or patio near ground level; (3) concave corner, where 2 short (≤5 m) façades united to form a small indentation in the building outline; (4) convex round, where the façade curved without any well-defined corners to form a protrusion in the building outline (note that a concave round would be considered an alcove); and (5) flat, where the façade formed a plane, perhaps with some small protrusions or indentations (typically ornamentations).
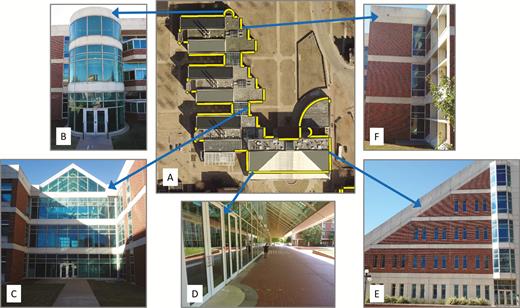
Façade type examples from one building at Oklahoma State University (OS03 – Noble Research Center) monitored for bird–window collisions 2015–2017. (A) Aerial view of one building showing façade search areas (yellow polygons); façade perimeters with no search area were not monitored because they lacked windows. Façade types (described in text) included (B) convex rounds, (C) alcoves, (D) porticos, (E) flats, and (F) concave corners.
We calculated the remaining variables (land cover and distance to trees) using spatial data layers provided by the department of Geospatial Systems at Oklahoma State University (OSUGS), which included georectified aerial imagery, points of individual tree bole locations, and polygons of some land cover types (buildings, parking lots, sidewalks, lawns, and flower beds). These data were limited to the main OSU campus, except for the aerial imagery, which covered the entire study area. For off-campus buildings, we used the aerial imagery (spatial resolution varied between 0.05 and 1.00 m, but was mostly <0.25 m) and ground-truthing to digitize locations of individual trees. For the entire study area, we used aerial imagery to digitize polygons for cover classes that, spatially or categorically, were not included in the OSUGS land cover layer. We aggregated land cover into 3 cover classes: impervious (e.g., asphalt and concrete), lawn (short, maintained turf grasses), and flowerbeds (including hedges and shrubs). Because most façade search areas lacked tree canopy coverage and tree canopy was unrelated to cover at ground level, we did not consider it when classifying land cover. We calculated the proportion of each 2-m-wide façade search area polygon overlapped by each cover class. We calculated distance to trees as the mean distance to the 3 trees nearest each façade polygon (trees within the polygon had a distance of 0 m). We used multiple trees, rather than only the nearest tree, to better characterize the proximity of tall vegetation generally.
Distance of building façades to the nearest body of water may also influence collision risk because water bodies and their associated shoreline vegetation influence avian habitat use, movements, abundance, and species composition. However, we did not consider this factor in analyses because (1) our study area contains no major rivers and only one sizeable lake of ~80 ha; and (2) 11 of our 16 study buildings (those on the OSU campus) were clustered in a ~60 ha area ≥2 km from this lake, which limits both the variation of distance to water among building façades in our study area and the likely importance of this factor in influencing collisions.
Statistical Analyses
We conducted statistical analyses in R 3.5.2 (R Core Team 2018) with RStudio 1.1.463 (RStudio Team 2016). Although we assumed that a carcass location corresponded to a collision at the nearest façade, the source and location of mortality for feather piles had greater uncertainty because a feather pile could represent a predator-killed bird or a window-killed bird that a scavenger moved away from the collision site before consuming. For fatal collision counts across all species, we therefore conducted analyses both including and excluding feather piles.
For all sets of analyses described below, we adjusted carcass counts to account for 2 major survey-related biases that cause underestimation of mortality: removal of carcasses between surveys by humans and animal scavengers, and imperfect detection of carcasses by surveyors. These adjustments were based on experimental removal and detection trials conducted at the same buildings for a companion study (Riding and Loss 2018). For removal trials, bird carcasses were placed in areas where collisions could occur, then monitored by trail cameras and daily surveyor checks until all observable remains were removed. For detection trials, a survey coordinator placed carcasses within building survey areas prior to a collision survey being conducted, and the rate at which surveyors detected these carcasses was documented. Detection trials were conducted for all authors and technicians that regularly conducted collision surveys; all surveyors were aware that a detection trial could occur at any time throughout the study but were blind to the specific date and location of each trial. Both removal and detection trials were conducted in all seasons during which collision monitoring occurred, and bird carcasses used for trials were obtained as collision casualties. Data from the removal and detection trials were used in the R package carcass (Korner-Nievergelt et al. 2015) to estimate daily carcass persistence probability and detection probability, which were then used in a statistical estimator (Korner-Nievergelt et al. 2013) to generate bias adjustment factors we applied to carcass counts in this study.
We treated individual façades as replicates to assess the importance of façade-level variables in explaining bias-adjusted carcass counts at each façade. Because response variables were continuous with a large number of zeros (i.e., we observed zero carcasses at many façades over the course of the entire study), we used a compound Poisson–gamma mixed model (function cpglmm in package cpglm; Zhang 2013), a type of Tweedie distribution model that handles continuous, zero-inflated data without treating the zero and non-zero values separately (Lecomte et al. 2013). We specified a random effect for building because façades nested within buildings were not independent of each other. Because numbers of surveys (even within the same building) often varied as a result of some buildings or façades being inaccessible due to construction or other activities, we standardized for effort by specifying an offset term for number of surveys at each façade (analyses for individual seasons included numbers of surveys for that season only). We conducted univariate correlation analyses among all possible pairs of numeric predictor variables. We did not use impervious cover and lawn cover together in the same model because they were strongly correlated (|r| > 0.70); however, these 2 variables did appear in different models in the same model selection exercise for each response variable.
When assessing bias-adjusted carcass counts for all species combined, 3 of 4 façades at building OS12 appeared to be outliers for spring, summer, and all seasons combined (Supplemental Material Figure S1), but not for fall or species-specific analyses. Therefore, we performed these analyses both including and excluding OS12 to assess effects on model selection results. We did this rather than simply excluding OS12 from analyses because it was responsible for 35% (n = 154) of all carcass observations across seasons, and its exclusion would have greatly reduced replication of collisions for our study. Ultimately, we assessed predictor variables against 16 total response variables: 3 response variables for total collisions combined across all seasons and species (1 including building OS12 and feather piles; 1 excluding OS12 and including feather piles; and 1 including OS12 and excluding feather piles); 5 response variables for collisions in separate seasons combined across all species (2 for spring, 1 including and 1 excluding building OS12; 2 for summer, 1 including and 1 excluding OS12; and 1 for fall, which included OS12); and 8 response variables for collisions of separate species combined across seasons (building OS12 included for all 8 analyses).
All response variables were bias-adjusted carcass counts, but for brevity, we hereafter refer to response variables as “collisions.” To derive important predictor variables for each response variable, we used a 3-step model selection procedure with Akaike’s information criterion corrected for small sample size (ΔAICc; Burnham and Anderson 2002). We used this 3-step process rather than automated approaches like stepwise model selection because those typically result in a single model, whereas we were interested in comparing among several potentially competitive models. First, we compared ΔAICc values for a null model and 8 single-variable models, each containing 1 of the 8 façade variables individually. The 3 top-ranked single-variable models (i.e., lowest ΔAICc) were used to construct 17 two-variable models, which collectively contained all possible additive combinations of variables from the top 3 single-variable models (unless impervious cover or lawn cover was in a top-ranked single-variable model, in which case there were only 15 or 16 two-variable models). Second, we compared ΔAICc values among the null model, top-ranked single variable models, and 15 to 17 two-variable models. The 3 top-ranked models from this second comparison were used in combination with other variables appearing in the 10 top-ranked models to make 3 sets of two- to five-variable additive models that had not already been assessed. Third, we compared ΔAICc values among the null model, the 5 top-ranked models from step 2, and 6 to 9 additional multiple variable models generated in step 2.
We interpreted models that were strongly supported (i.e. ΔAICc < 2) in the third model selection step only. When multiple models had strong support in this final model selection step, we combined them for interpretation using conditional model averaging (function model.avg in R package MuMIn). We did not consider any models for interpretation that were more complex versions of higher-ranking nested models (i.e. models that contained uninformative variables; Richards 2008, Arnold 2010). For the single categorical variable (façade type), the baseline category used for comparisons was “alcove.” We considered estimated coefficients of other categories to be meaningfully different if the standard error (SE) was less than the absolute value of the estimated coefficient (Stephens et al. 2005).
Finally, façade type could not be included in fall and species-specific analyses (with one exception) because at least one façade type experienced no collisions, which caused ill-defined coefficient estimates with SEs that were orders of magnitude larger. We therefore dropped levels of façade type that lacked collisions to better assess whether this variable was an important correlate of collisions. If façade type was found to be unimportant, we retained and interpreted results of the original analysis that excluded the façade type variable. If façade type was important, we interpreted results of the analysis with façade type. We report the levels of façade type dropped, and the subsequent decrease in number of façade replicates (Supplemental Material Tables S4–S11).
RESULTS
From April 2015 to May 2017, we conducted 6,190 total building surveys (2,270 spring, 2,340 summer, 1,580 fall) and recorded 63 bird species as casualties of window collisions, including 418 carcasses (214 spring, 104 summer, 100 fall) of which 71 were feather piles (25 spring, 35 summer, 11 fall). Among 235 façades at 16 buildings (range: 4–41 façades per building), we recorded collisions at 88 façades across 14 buildings (spring: 57 façades at 13 buildings; summer: 41 façades at 13 buildings; fall: 38 façades at 11 buildings).
Barring 2 exceptions, model selection results including and excluding the likely outlier building (OS12) were nearly identical, with models ranked in the same relative order and estimated coefficients having the same sign and very minor estimated differences in effect sizes. The first exception was for the analyses of both total collisions across all species and seasons and of spring collisions across all species, where excluding OS12 caused the convex round façade type to have a meaningful negative effect size. In other words, convex rounds experienced fewer collisions compared to alcoves when we excluded OS12 but not when we included OS12. The second exception was for the analysis of summer collisions across all species, where façade length was present in the strongly supported model when we included OS12 but absent from the strongly supported model when we excluded OS12. For analyses of total collisions across all seasons and species, results including and excluding feather piles were likewise similar. Further, because most feather piles could not be identified to species, they were rarely included in species-specific analyses. Because we have noted these relatively slight differences here, results presented and discussed below are based on analyses that include both OS12 and feather piles.
Total Collision Mortality
For collision mortality across seasons and species, only one model received strong support (Table 1). This model indicated that, among façade types, alcoves experienced more collision mortality than did corners (β = −1.07, SE = 0.50), flats (β = −0.99, SE = 0.27), and porticos (β = −0.97, SE = 0.42) but did not differ meaningfully from convex rounds (β = −0.23, SE = 0.42; see above exception when OS12 was excluded). Also, total collisions were positively associated with glass proportion (β = 3.01, SE = 0.41), façade height (β = 0.10, SE = 0.02), and façade length (β = 0.02, SE = 0.01). Notably, these variables all represented aspects of façade size or structure, and no vegetation or land cover variables were represented in the top model for bird collision mortality.
Akaike’s information criterion (ΔAICc) model ranking for analysis of relationships between façade variables and bias-adjusted counts of total fatal bird collisions across all seasons and species, based on bird–window collision monitoring in Stillwater, Oklahoma, 2015–2017. Number of parameters (k) and model weights (wi) are also given. Potential predictor variables include proportion of façade surface covered by glass (Glass), façade height (Height), façade length (Length), façade type (Type), mean tree distance (Tree), and land cover by flowerbeds (Flower), impervious surfaces (Imperv), and lawns (Lawn).
| Variables | ΔAICc | k | wi |
|---|---|---|---|
| Glass, Height, Length, Type | 0.0 | 10 | 0.940 |
| Glass, Height, Length | 5.6 | 6 | 0.058 |
| Glass, Height | 14.2 | 5 | <0.001 |
| Glass, Length, Type | 15.3 | 9 | <0.001 |
| Glass, Length | 29.6 | 5 | <0.001 |
| Glass, Type | 37.0 | 8 | <0.001 |
| Glass | 43.1 | 4 | <0.001 |
| Null | 72.6 | 3 | <0.001 |
| Variables | ΔAICc | k | wi |
|---|---|---|---|
| Glass, Height, Length, Type | 0.0 | 10 | 0.940 |
| Glass, Height, Length | 5.6 | 6 | 0.058 |
| Glass, Height | 14.2 | 5 | <0.001 |
| Glass, Length, Type | 15.3 | 9 | <0.001 |
| Glass, Length | 29.6 | 5 | <0.001 |
| Glass, Type | 37.0 | 8 | <0.001 |
| Glass | 43.1 | 4 | <0.001 |
| Null | 72.6 | 3 | <0.001 |
Akaike’s information criterion (ΔAICc) model ranking for analysis of relationships between façade variables and bias-adjusted counts of total fatal bird collisions across all seasons and species, based on bird–window collision monitoring in Stillwater, Oklahoma, 2015–2017. Number of parameters (k) and model weights (wi) are also given. Potential predictor variables include proportion of façade surface covered by glass (Glass), façade height (Height), façade length (Length), façade type (Type), mean tree distance (Tree), and land cover by flowerbeds (Flower), impervious surfaces (Imperv), and lawns (Lawn).
| Variables | ΔAICc | k | wi |
|---|---|---|---|
| Glass, Height, Length, Type | 0.0 | 10 | 0.940 |
| Glass, Height, Length | 5.6 | 6 | 0.058 |
| Glass, Height | 14.2 | 5 | <0.001 |
| Glass, Length, Type | 15.3 | 9 | <0.001 |
| Glass, Length | 29.6 | 5 | <0.001 |
| Glass, Type | 37.0 | 8 | <0.001 |
| Glass | 43.1 | 4 | <0.001 |
| Null | 72.6 | 3 | <0.001 |
| Variables | ΔAICc | k | wi |
|---|---|---|---|
| Glass, Height, Length, Type | 0.0 | 10 | 0.940 |
| Glass, Height, Length | 5.6 | 6 | 0.058 |
| Glass, Height | 14.2 | 5 | <0.001 |
| Glass, Length, Type | 15.3 | 9 | <0.001 |
| Glass, Length | 29.6 | 5 | <0.001 |
| Glass, Type | 37.0 | 8 | <0.001 |
| Glass | 43.1 | 4 | <0.001 |
| Null | 72.6 | 3 | <0.001 |
Seasonal Collision Mortality
For spring collisions, only one model received strong support (Supplemental Material Table S1), and it included the same variables (with similar coefficients and SEs) as the top model for total collisions (i.e. façade proportion glass, height, length, and type). Given that we observed more fatal collisions in spring than in summer and fall combined, it follows that spring results closely mirror results for all seasons combined. For summer collisions, there was only one strongly supported model (Supplemental Material Table S2), which differed slightly from the top total collision model in having façade type replaced by lawn cover as an important variable. Summer collisions were positively associated with glass proportion (β = 2.76, SE = 0.49), façade height (β = 0.11, SE = 0.02), façade length (β = 0.01, SE < 0.01), and lawn cover (β = 0.86, SE = 0.29). For fall collisions, there was one strongly supported model (Supplemental Material Table S3), with collisions positively related to glass proportion (β = 3.51, SE = 0.64), façade height (β = 0.08, SE = 0.04), mean tree distance (β = 0.04, SE = 0.01), and lawn cover (β = 0.66, SE = 0.42). Among façade types (except for corners, which were dropped because of zero collisions), alcoves experienced more fall collisions than flats (β = −0.45, SE = 0.40) but did not differ meaningfully from porticos (β = −0.16, SE = 0.53) or convex rounds (β = −1.10, SE = 1.13). In summary (Table 2), structural aspects of a façade (especially height and proportion covered by glass) seemed more strongly correlated with collisions for separate seasons than did vegetation and land cover variables.
Comparison of relationships for variables from strongly supported models for 4 different collision response variables (bias-adjusted fatal collision counts across all species and seasons, and for spring, summer, and fall) based on bird–window collision monitoring in Stillwater, Oklahoma, 2015–2017. Potential predictor variables include proportion of façade surface covered by glass (Glass), façade height (Height), façade length (Length), façade type (Type), mean tree distance (Tree), and land cover by flowerbeds (Flower), impervious surfaces (Imperv), and lawn (Lawn). Positive and negative coefficients for predictor variables included in top models are indicated by + and −, respectively; predictor variables not appearing in top models are indicated by 0. When more than one model was strongly supported, inference was based on averaged coefficients. For façade type, the type with the highest mortality is indicated.
| Response variable | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| Multi-season | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Spring | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Summer | 1 | + | + | + | 0 | 0 | 0 | 0 | + |
| Fall | 1 | + | + | 0 | multiple | + | 0 | 0 | + |
| Response variable | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| Multi-season | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Spring | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Summer | 1 | + | + | + | 0 | 0 | 0 | 0 | + |
| Fall | 1 | + | + | 0 | multiple | + | 0 | 0 | + |
Comparison of relationships for variables from strongly supported models for 4 different collision response variables (bias-adjusted fatal collision counts across all species and seasons, and for spring, summer, and fall) based on bird–window collision monitoring in Stillwater, Oklahoma, 2015–2017. Potential predictor variables include proportion of façade surface covered by glass (Glass), façade height (Height), façade length (Length), façade type (Type), mean tree distance (Tree), and land cover by flowerbeds (Flower), impervious surfaces (Imperv), and lawn (Lawn). Positive and negative coefficients for predictor variables included in top models are indicated by + and −, respectively; predictor variables not appearing in top models are indicated by 0. When more than one model was strongly supported, inference was based on averaged coefficients. For façade type, the type with the highest mortality is indicated.
| Response variable | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| Multi-season | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Spring | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Summer | 1 | + | + | + | 0 | 0 | 0 | 0 | + |
| Fall | 1 | + | + | 0 | multiple | + | 0 | 0 | + |
| Response variable | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| Multi-season | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Spring | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Summer | 1 | + | + | + | 0 | 0 | 0 | 0 | + |
| Fall | 1 | + | + | 0 | multiple | + | 0 | 0 | + |
Species-Specific Collision Mortality
In decreasing order of frequency and representing 47% of our total collisions, the 8 species with sufficient observations (n ≥ 15) of collision fatalities to analyze individually were Lincoln’s Sparrow (Melospiza lincolnii; n = 41), Mourning Dove (Zenaida macroura; n = 29), Swainson’s Thrush (Catharus ustulatus; n = 28), European Starling (Sturnus vulgaris; n = 27), House Finch (Haemorhous mexicanus; n = 21), Painted Bunting (Passerina ciris; n = 19), American Robin (Turdus migratorius; n = 17), and Indigo Bunting (Passerina cyanea; n = 15). Overall, species-specific analyses showed more variation in collision correlates than did season-specific analyses, with vegetation and land cover variables appearing in more supported models (Table 3).
Comparison of relationships for variables from strongly supported models for 9 different collision response variables (bias-adjusted fatal collision counts across all species and for 8 individual species) based on bird–window collision monitoring in Stillwater, Oklahoma, 2015–2017. Potential predictor variables include proportion of façade surface covered by glass (Glass), façade height (Height), façade length (Length), façade type (Type), mean tree distance (Tree), and land cover by flowerbeds (Flower), impervious surfaces (Imperv), and lawn (Lawn). Positive and negative coefficients for predictor variables included in top models are indicated by + and −, respectively; predictor variables not appearing in top models are indicated by 0. When more than one model was strongly supported, inference was based on averaged coefficients. For façade type, the type with the highest mortality is indicated.
| Species | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| All species | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Lincoln’s Sparrow | 2 | + | 0 | 0 | 0 | + | − | 0 | 0 |
| Mourning Dove | 1 | + | + | 0 | 0 | 0 | 0 | 0 | 0 |
| Swainson’s Thrush | 2 | + | + | + | round | 0 | 0 | 0 | 0 |
| European Starling | 1 | 0 | 0 | 0 | multiple | 0 | − | 0 | 0 |
| House Finch | 3 | + | + | + | round | + | 0 | − | 0 |
| Painted Bunting | 2 | + | + | 0 | multiple | 0 | 0 | + | 0 |
| American Robin | 2 | 0 | 0 | + | 0 | 0 | 0 | − | + |
| Indigo Bunting | 2 | 0 | + | + | alcove | 0 | 0 | 0 | 0 |
| Species | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| All species | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Lincoln’s Sparrow | 2 | + | 0 | 0 | 0 | + | − | 0 | 0 |
| Mourning Dove | 1 | + | + | 0 | 0 | 0 | 0 | 0 | 0 |
| Swainson’s Thrush | 2 | + | + | + | round | 0 | 0 | 0 | 0 |
| European Starling | 1 | 0 | 0 | 0 | multiple | 0 | − | 0 | 0 |
| House Finch | 3 | + | + | + | round | + | 0 | − | 0 |
| Painted Bunting | 2 | + | + | 0 | multiple | 0 | 0 | + | 0 |
| American Robin | 2 | 0 | 0 | + | 0 | 0 | 0 | − | + |
| Indigo Bunting | 2 | 0 | + | + | alcove | 0 | 0 | 0 | 0 |
Comparison of relationships for variables from strongly supported models for 9 different collision response variables (bias-adjusted fatal collision counts across all species and for 8 individual species) based on bird–window collision monitoring in Stillwater, Oklahoma, 2015–2017. Potential predictor variables include proportion of façade surface covered by glass (Glass), façade height (Height), façade length (Length), façade type (Type), mean tree distance (Tree), and land cover by flowerbeds (Flower), impervious surfaces (Imperv), and lawn (Lawn). Positive and negative coefficients for predictor variables included in top models are indicated by + and −, respectively; predictor variables not appearing in top models are indicated by 0. When more than one model was strongly supported, inference was based on averaged coefficients. For façade type, the type with the highest mortality is indicated.
| Species | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| All species | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Lincoln’s Sparrow | 2 | + | 0 | 0 | 0 | + | − | 0 | 0 |
| Mourning Dove | 1 | + | + | 0 | 0 | 0 | 0 | 0 | 0 |
| Swainson’s Thrush | 2 | + | + | + | round | 0 | 0 | 0 | 0 |
| European Starling | 1 | 0 | 0 | 0 | multiple | 0 | − | 0 | 0 |
| House Finch | 3 | + | + | + | round | + | 0 | − | 0 |
| Painted Bunting | 2 | + | + | 0 | multiple | 0 | 0 | + | 0 |
| American Robin | 2 | 0 | 0 | + | 0 | 0 | 0 | − | + |
| Indigo Bunting | 2 | 0 | + | + | alcove | 0 | 0 | 0 | 0 |
| Species | Models | Glass | Height | Length | Type | Tree | Flower | Imperv | Lawn |
|---|---|---|---|---|---|---|---|---|---|
| All species | 1 | + | + | + | alcove | 0 | 0 | 0 | 0 |
| Lincoln’s Sparrow | 2 | + | 0 | 0 | 0 | + | − | 0 | 0 |
| Mourning Dove | 1 | + | + | 0 | 0 | 0 | 0 | 0 | 0 |
| Swainson’s Thrush | 2 | + | + | + | round | 0 | 0 | 0 | 0 |
| European Starling | 1 | 0 | 0 | 0 | multiple | 0 | − | 0 | 0 |
| House Finch | 3 | + | + | + | round | + | 0 | − | 0 |
| Painted Bunting | 2 | + | + | 0 | multiple | 0 | 0 | + | 0 |
| American Robin | 2 | 0 | 0 | + | 0 | 0 | 0 | − | + |
| Indigo Bunting | 2 | 0 | + | + | alcove | 0 | 0 | 0 | 0 |
For Lincoln’s Sparrow, there were 2 strongly supported models (Supplemental Material Table S4) that indicated collisions increased with increasing proportion of glass (β = 3.04, SE = 0.61) and mean tree distance (β = 0.02, SE = 0.01) and with decreasing cover of flowerbeds (β = −2.10, SE = 1.78). This was 1 of 3 species for which façade height was not in a strongly supported model. The other 2 such species, American Robin and European Starling, were also 2 of 3 species for which glass proportion was not in a strongly supported model. The robin had 2 strongly supported models (Supplemental Material Table S5) indicating collisions increased with increasing façade length (β = 0.03, SE = 0.01) and lawn cover (β = 1.42, SE = 0.70) and decreasing impervious cover (β = −1.49, SE = 0.71). The starling had one strongly supported model (Supplemental Material A Table S6) wherein collisions increased with decreasing flowerbed cover (β = −11.42, SE = 10.37) and occurred less frequently at alcoves than at flats (β = 2.25, SE = 1.04) and convex rounds (β = 2.25, SE = 1.06). The starling was the only species for which alcoves had fewer collisions than other analyzed façade types, but both corners and porticos were dropped from analysis for having zero collision observations, indicating alcoves did not experience the fewest starling collisions.
The only species for which all façade types could be analyzed was Mourning Dove, but façade type did not appear in the single strongly supported model for this species (Supplemental Material A Table S7). Mourning Dove collisions were positively related to façade height (β = 0.17, SE = 0.06) and proportion of glass (β = 3.11, SE = 1.17). Façade type was in at least one strongly supported model for all other species (Supplemental Material A Tables S8–S11). For Indigo Bunting, alcoves had more collisions than did corners (β = −1.65, SE = 1.12), flats (β = −5.03, SE = 1.77), and convex rounds (β = −2.53, SE = 1.24). Convex rounds were the only façade type to have more collisions than alcoves for Swainson’s Thrush (β = 1.29, SE = 0.58) and House Finch (β = 2.50, SE = 1.21). For Painted Bunting, flats had fewer collisions (β = −1.59, SE = 0.58), but the number of collisions at porticos (β = −0.01, SE = 0.89) and convex rounds (β = 0.06, SE = 1.07) was not meaningfully different from alcoves.
For Indigo Bunting and Swainson’s Thrush, each with 2 strongly supported models, there was no apparent effect of any vegetation or land cover variables; however, façade structural variables were associated with collisions. Indigo Bunting collisions were positively related with façade height (β = 0.09, SE = 0.06) and length (β = 0.09, SE = 0.03), while Swainson’s Thrush collisions were positively related with façade height (β = 0.17, SE = 0.05), length (β = 0.02, SE = 0.01), and glass proportion (β = 5.58, SE = 1.12).
Collisions for the remaining 2 species were related to façade structural features and vegetation or cover variables. Based on 3 strongly supported models, House Finch collisions were positively associated with façade height (β = 0.23, SE = 0.09), façade length (β = 0.04, SE = 0.02), glass proportion (β = 4.93, SE = 2.21), and mean tree distance (β = 0.09, SE = 0.06), and negatively associated with impervious cover (β = −3.37, SE = 1.42). From 2 strongly supported models, Painted Bunting collisions were positively related with façade height (β = 0.22, SE = 0.05), glass proportion (β = 3.16, SE = 1.21), and impervious cover (β = 1.50, SE = 0.74).
Discussion
Important Façade-Level Predictors of Collision Mortality
We assessed the influence of fine-scale (i.e. façade-level) building structural, vegetation, and land cover–related variables on fatal bird–window collisions in a small urban area of the central United States to inform collision deterrence practices that could be targeted within individual buildings. The proportion of glass on the façade surface was the predictor variable supported for the greatest number of collision response variables, including for total collisions across all species and seasons, across all species for spring, summer, and fall, and across all seasons for 5 of the 8 species analyzed separately. In other words, more collisions occurred at façades with a larger proportion of glass, a general result that corroborates previous studies wherein the amount or proportion of glass across entire buildings positively correlated with collisions (e.g., Klem et al. 2009, Borden et al. 2010, Hager et al. 2013, Keyes and Sexton 2014, Cusa et al. 2015, Barton et al. 2017, Schneider et al. 2018). However, this is the first confirmation that such a relationship also occurs at the scale of individual building façades.
Although extensive glass surfaces certainly contribute to elevated bird collision risk, conflating factors that we did not account for may have contributed somewhat to the apparent importance of glass proportion in our study. For example, glass proportion is likely correlated with both the proximity and contiguousness of glass areas; in other words, windows tend to be closer together and glass has larger contiguous area when glass proportion is greater, and these properties may make it more difficult for birds to perceive the glass. Also, glass proportion is likely to be positively correlated with the amount and/or intensity of light emitted at night (i.e. more light emitted from façades with more window area; Parkins et al. 2015). This light pollution may attract and confuse nocturnally migrating birds, increasing collisions at buildings (Evans Ogden 2002, Gauthreaux and Belser 2006, Haupt and Schillemeit 2011, Keyes and Sexton 2014, Parkins et al. 2015, Winger et al. 2019).
Next to glass proportion, the most frequently supported variables represented aspects of façade size (i.e. height and length). This follows previous studies in finding more collisions or higher collision risk at larger buildings (Machtans et al. 2013, Loss et al. 2014, Hager et al. 2017), but again, this finding had yet to be confirmed at the finer façade scale. Large façades provide more area for collisions relative to smaller façades with the same proportion of glass. In combination with the overwhelming importance of glass proportion, our results therefore suggest that large façades with high proportions of glass pose the greatest collision risk to birds. Thus, collision deterrence efforts may need to focus on large façades independent of overall building size.
In addition to the structural variables discussed above, façade type also appeared in supported models for several collision response variables. For collisions across all species, including both across all seasons and separately for spring, there was greater mortality at alcoves than other façade types. Building and façade shape have never been formally assessed in relation to window collisions, and this finding could indicate that the tunnel-like nature of alcoves, especially deep alcoves with tall façades, may “trap” birds (although responses appear to vary somewhat among species; Table 3). Such entrapment may be exacerbated by unique lighting, reflection, and/or see-through effects that occur in deeper alcoves with decreased penetration by natural light, and trapped birds may be at greater risk of subsequent collision if the end of the alcove has a large proportion of glass that appears to provide a safe exit. Porticos are similarly tunnel-like but had much lower collision rates overall. The relatively low mortality at porticos may arise from birds more easily recognizing the horizontal exit paths associated with this façade type as opposed to the vertical exit path associated with alcoves, and this differential perception may in turn be related to birds in flight generally having larger visual fields laterally than above the head (Martin 2012).
Equally notable is that vegetation and land cover variables did not appear in the top model for total collisions across all seasons and species. This result was somewhat unexpected given that such variables have previously been correlated with collisions at broader scales (Klem et al. 2009, Borden et al. 2010, Hager et al. 2013, Kummer et al. 2016, Elmhurst and Grady 2017) and at other types of glass structures (Sierro and Schmid 2001, Barton et al. 2017). However, these variables were not entirely unimportant, as they appeared in top models for several season- and species-specific analyses. In fact, the lack of support for such variables in explaining total bird collisions may partially arise from opposing season- or species-specific effects (e.g., opposite relationships of House Finch and Painted Bunting collisions with impervious cover) that cancel each other out when seasons and species are aggregated. This potential lack of influence on total collisions by vegetation and land cover variables at the façade scale is certainly an area that requires further investigation; specifically, studies should be conducted in other regions and should assess other vegetation and land cover types to determine if and when these factors affect total collisions at the façade scale.
Seasonal Collision Mortality
Analyses of collisions for separate seasons generated results that differed slightly from the analysis of total collisions across seasons, except for the spring analysis which documented the exact same collision correlates as the total collision analysis, which is expected given that the greatest number of collisions occurred during spring. Summer and fall analyses did not support the predictive importance of some structural variables (façade length and type) that were supported in the total collision analysis; however, additional collision correlates were also revealed for these seasons (lawn cover and tree distance). These results indicate that collision correlates differ seasonally, a finding that has only been documented once for the issue of bird–window collisions (Loss et al. 2019), and which suggests that results of past studies that analyzed data across all seasons or from a single season only should not necessarily be extrapolated to individual seasons. Fall migration is often assumed to be the most critical period for window collisions, because collision frequency or mortality is often reported to be greater in fall than any other season (Zink and Eckles 2010, Kummer and Bayne 2015, Bracey et al. 2016). However, collision mortality in seasons other than fall may be underappreciated, and may in some cases even exceed fall mortality (Dunn 1993, Gómez-Moreno et al. 2018, Schneider et al. 2018, Brown et al. 2019). Thus, identification and consideration of season-specific collision correlates will be important to consider in areas where substantial mortality occurs in seasons other than fall.
As seasonal assessment of bird–building collision correlates is novel, we list 2 caveats related to our study that could limit the generalizability of these seasonal findings. First, results from this same study area, and based on the same collision data, suggest that patterns of temporal variation in collisions likely differ geographically. Specifically, our study area experiences an unusually high number of bird–building collisions during spring. Many of these spring collisions are of nonmigratory resident bird species; when solely considering migrating individuals (i.e. individuals of migratory species that collide during their migratory passage period in our study area), seasonal patterns more closely match previous studies, with more migrants colliding in fall than spring (C. R. Riding personal communication). Given the relatively unique temporal patterns of collisions, the factors driving façade-level variation in collisions may also be unique to our study area, and caution should be exercised when extrapolating season-specific results (as well as our results for total collision and species-specific analyses) to other geographic regions or study areas that are urbanized to a greater or lesser degree. Second, seasonal results may be confounded by species-specific results because our study area experiences substantial seasonal variation in the species composition of collision casualties (C. R. Riding personal communication). That is, these seasonal results may be at least partly influenced by which species were present (and colliding) in addition to seasonal changes that influence collision risk factors (e.g., compass bearing of the sun at sunrise, vegetation growth, availability of plant and insect food sources near buildings, and patterns of when and how frequently buildings are illuminated with artificial night lighting). Regardless of these caveats, we hypothesize that seasonal variation in collision correlates is widespread, even if the nature of such seasonal variations differs from those observed in this study.
Species-Specific Collision Mortality
Species-specific analyses showed highly variable collision correlates. For example, although at least one façade structure variable (e.g., glass proportion or façade height) appeared in the strongly supported models for every species, no variable was supported in top models for >5 of the 8 species assessed. In addition to the above-described importance of glass proportion, façade type was also important for 5 of 8 species, but the specific façade type(s) causing the most collisions varied among species. Moreover, for impervious cover, the direction of the effect differed among species (negative for American Robin and House Finch, positive for Painted Bunting). Although these 8 species represent nearly half of the collision fatalities we observed, they comprise only 13% of species observed as collision casualties.
Even pairs of species with relatively close taxonomic relationships (American Robin and Swainson’s Thrush; Painted Bunting and Indigo Bunting) had differing results, which suggests that factors other than those related to taxonomy are likely to influence collisions and that it may be difficult to extrapolate findings from one species to another. These species-varying factors could include, for example, migration and dispersal timing and length (e.g., Swainson’s Thrush is a nocturnal migrant while American Robin is primarily a diurnal migrant, and post-breeding dispersal and fall migration begin earlier in the season for Painted Bunting than Indigo Bunting) and foraging and habitat selection strategy (e.g., differential preference/avoidance of ornamental vegetation and lawns near buildings). Like the season-specific analyses, our finding of among-species variation in collision correlates indicates that analyses including all species may overlook important species-specific collision risk factors; however, the same caveat regarding potential conflating between season- and species-specific patterns also exists for this analysis.
Only 2 previous studies, both of which focused on the building scale, attempted to assess structural and vegetation correlates of bird–window collisions for individual species. In Cusa et al. (2015), species-specific results were aggregated into feeding and habitat guilds for ease of interpretation; this analysis showed, for example, that foliage gleaners from forested habitats were more likely to collide at buildings surrounded by a greater extent of urban greenery. While we did not formally group species by life history traits (e.g., feeding guild, migratory strategy), those that we assessed (1 species in Columbiformes: Family Columbidae; 7 species in Passeriformes: Families Turdidae, Sturnidae, Passerellidae, Fringillidae, and Cardinalidae) represented modest diversity in taxonomy, as well as various life history strategies for diet, breeding habitat, urban adaptivity, and foraging and migration strategies. In Loss et al. (2019), window collision correlates were analyzed for 5 separate migratory bird species, none of which we assessed here. That study found that glass area and/or proportion of surrounding land covered by vegetation were positively associated with collisions for all 5 species, but there were also some differences in correlates among species (e.g., building footprint and height positively associated with collisions for only one species, Nashville Warbler [Leiothlypis ruficapilla]; no association between glass area and collisions for one species, Tennessee Warbler [Leiothlypis peregrina], and an association between collisions and amount of vegetation within 100 m for Common Yellowthroat, whereas collisions for all other species were related to vegetation within 50 m).
The above life history variation that we captured across the 8 species assessed likely explains some of the among-species differences in collision correlates. The most disparate species-level results were for American Robin and Painted Bunting, which shared zero (of a possible 8) strongly supported variables with the same direction of relationship. Robin collisions were negatively related to impervious cover (e.g., concrete) and positively related to lawn cover—although this finding likely arose at least partly due to the strong inverse correlation between these 2 predictor variables—whereas for Painted Bunting, collisions were positively related to impervious cover and there was no support for an effect of lawn cover. These differences may be related to varying foraging strategies and/or the types of activity birds were engaged in prior to colliding. Robins frequently forage on lawns and adapt well to urban settings, making them likely to forage on a lawn (but not on concrete) near a building. If startled into flight by a perceived predator (e.g., human or dog [Canis lupus familiaris]), a robin may occasionally flee toward a building, perhaps even directly at a window if it is misperceived as an opening in the building (Ros et al. 2017). Painted Buntings often forage on the ground (albeit in taller grasses than those used in lawns), but are likely to be present in highly urbanized areas during migration or dispersal only. The positive relationship between bunting collisions and impervious cover may arise if buntings become confused (e.g., in alcoves—see above) when they stop in highly developed areas during migration. In order to enact species-specific management, further research may be needed to clarify mechanisms of the collision process related to different bird species and life history traits.
Limitations and Future Research
In addition to considering the above-mentioned caveats, our results should be interpreted with caution because the large number of analyses with assessment of variable importance based on standard errors of coefficient estimates increases the probability of apparently meaningful effects arising by chance (Type I error). Further, although our results for some collision correlates (e.g., glass proportion) were robust to the choice of data analyzed, indicating the importance of these factors in influencing bird collisions, results for some analyses depended on whether we included one exceptionally glassy, outlier building (OS12). Specifically, for analyses of total collisions across seasons and for spring only, round-shaped façades experienced fewer collisions only when OS12 was excluded, and for the summer collision analysis, façade length was only important when this building was included. Thus, our findings related to these potential collision correlates are not necessarily broadly generalizable beyond our study area.
This study only assessed façade-level variation in bird collisions; however, the effects of façade-related features are also likely mediated by broader-scale factors, including those operating at the scales of entire buildings and of the surrounding landscape and region (Hager et al. 2017). Additionally, we hypothesize that some factors may be important at one scale but not another (e.g., an association between bird collisions and a feature measured at the façade level, but no association with the same feature measured at the building level), and that the scale at which collision risk factors operate may vary seasonally and/or by bird species due to temporal variation in resource availability and taxonomic variation in the scale of habitat selection. This expectation is supported by the above-mentioned result from Loss et al. (2019), where collisions of Common Yellowthroat were associated with the amount of vegetation within 100 m of buildings while collisions of 4 other species were associated with vegetation within 50 m. Future conservation efforts would benefit from research integrating risk factors at multiple scales into a unified understanding of spatiotemporal factors influencing bird–window collisions.
Conclusions
Our novel façade-level results, along with past research focusing on bird collision correlates at building, landscape, and regional scales, are informative for efforts to make buildings more bird-friendly by refining collision deterrence efforts and architectural and landscape designs. At the façade level, bird–window collisions seem to be most strongly driven by the proportion of glass covering a façade. Façade height and length also positively influenced number of collisions, and alcove-shaped façades caused more total collisions than any other façade type. Therefore, collision deterrence efforts should be targeted toward large and/or alcove-shaped façades covered by a large proportion of glass, and building designers should consider reducing and/or avoiding the use of such design features whenever possible.
Several conservation organizations have published guidelines to reduce bird collision risk at existing and newly constructed buildings (U.S. Fish and Wildlife Service 2016, American Bird Conservancy 2019, Fatal Light Awareness Program [FLAP] Canada 2019). Elements of these guidelines have been integrated into green building certification programs (e.g., the LEED program; U.S. Green Building Council 2016) and adopted and proposed legislation for bird-friendly building construction and retrofitting (e.g., San Francisco Planning Department 2019, U.S. Congress 2019). Although these guidelines and requirements recognize that collision risk varies within individual buildings, they are largely based on research that assessed collision risk for all birds and seasons combined, and they primarily include recommendations at the scale of entire buildings. Our study generally supports building-level research and collision mitigation guidelines that indicate bird collisions increase with more glass and greater structure size. However, our façade-level analysis provides additional nuance and quantitative support for focusing management on the building façades that are most problematic for birds. Further, our species- and season-specific assessments of collision correlates indicate that management guidelines based on studies assessing collision risk for all birds combined may not be equally effective in reducing collisions in all seasons and for all species. Future research, policy, and management that integrates information about collision risk for all bird species and seasons, and at multiple scales of variation from individual building façades to the surrounding landscape, should be most effective at reducing total bird–building collisions and thus benefiting avian conservation efforts.
Acknowledgments
We thank the building owners and managers that permitted us to conduct collision surveys, including OSU Facilities Management, Stillwater Designs, and CStar Management. We thank C. Barton, K. Emerson, C. McKinney, and >50 volunteer scientists for conducting carcass surveys. We thank I. Washburn for statistical consultation and B. Hilson and OSU Geospatial Systems for providing GIS data. We are grateful to S. Cady, J. Elmore, S. Lao, Y. Malyutina, and others for providing feedback that improved the manuscript.
Funding statement: This work was supported by the Oklahoma Agricultural Experiment Station, the Oklahoma State University Department of Natural Resource Ecology and Management, and by the National Institute of Food and Agriculture (United States Department of Agriculture) Hatch Grant Funds through the Oklahoma Agricultural Experiment Station (OKL-02915).
Ethics statement: We collected carcasses under federal (permit #MB05120C-0) and state (multiple permits over the study) scientific collecting permits, and protocols were approved by the OSU Institutional Animal Care and Use Committee (Animal Care and Use Protocol #AG-14–8).
Author contributions: All authors contributed to ideas, methodology design, resources, funding, data collection, editing, and revision. CSR conducted data analyses and writing.
Conflict of interest statement: We declare no conflict of interest.
Data depository: Analyses reported in this article can be reproduced using the data provided by Riding et al. (2020).

{kind=link}