





We have recently reported that pretreatment with electroacupuncture (EA) at stomach meridian point 36 (St36) prevents the chronic cold-stress increase in the hypothalamus-pituitary-adrenal axis (HPA), an action that may be under central control. Given that treatment for stress-related symptoms usually begins after onset of the stress responses, the objectives of the present study were to determine the efficacy of EA St36 on HPA hormones when EA St36 is given after stress was initiated, if the results are long lasting, and if blocking the glucocorticoid receptor (GR) using RU-486 had the same effects as EA St36. Adult male rats were placed in 4 groups of animals, 3 of which were exposed to cold and 1 of which was a nontreatment control group. After exposure to the cold stress, 2 groups were treated with either EA St36 or sham-EA, repeated over 10 days. The increase in ACTH and corticosterone observed in stress-only rats was prevented in EA St36 animals, and the effects remained intact 4 days after withdrawal of EA but continuation of cold stress. When the GR was blocked with RU-486, the efficacy of EA St36 remained unchanged. GR blockade did significantly elevate ACTH, which is not seen with EA St36, suggesting that EA St36 does act centrally. The elevated HPA hormones in stress-only rats were associated with a significant increase in depressive and anxious behavior; this was not observed in the stressed EA St36 animals. The results indicate that EA specifically at St36 vs sham-EA is effective in treating chronic poststress exposure.
Chronic stress has been directly linked to many disease states, affecting the health of numerous populations (1–3). Stress-induced perturbations occur through multiple biochemical and signaling pathways, with major involvement by the hypothalamus-pituitary-adrenal axis (HPA) and the sympathetic nervous systems (SNSs). Therefore, several treatment modalities for chronic stress have been directed at the HPA and SNS. Pharmaceutical therapies use drugs that either directly or indirectly affect these pathways, and include the anxiolytic benzodiazepines, the serotonin enhancing agents, and monoamine oxidase inhibitors (4). Moreover, lifestyle changes, such as yoga and meditation, have shown promise at abating the stress-induced HPA and SNS responses (5). Although these interventions are promising, no optimal treatment exists for chronic stress, and finding effective therapies for chronic stress-related disorders remains a major public health priority (6, 7).
Traditional Chinese medicine (TCM) acupuncture has been used for over 2500 years to treat various disorders, including stress. In recent decades the advent of electroacupuncture (EA), which is the application of electrical current via the inserted acupuncture needles, has added the benefit of delivering more accurate and uniform stimulation to several acupuncture points simultaneously (8). We have recently demonstrated that pretreatment with EA prevents the peripheral and central increases in HPA hormones caused by cold, painful stress over 10 days in a rat model (9). Specifically, stress-induced circulating ACTH and corticosterone (CORT), as well as central CRH mRNA and protein were blunted when EA at acupoint stomach meridian point 36 (St36) was performed before exposure to chronic cold stress (9). This strongly suggests that EA at St36 may be a viable therapeutic modality for chronic stress. A key issue, however, was whether the EA intervention would be effective when stress was already present.
In the current study, we tested the hypothesis that EA at St36 would be just as effective and as long lasting if delivered during concurrent exposure to chronic stress to better mimic how acupuncture is used clinically (10–12). Further, to confirm EA St36 activity on stress-induced HPA hormone release, in a separate experiment, we blocked glucocorticoid receptors (GRs) using the GR blocker, RU-486, and compared effects on the HPA with and without EA. In this cohort, we also examined the ability of EA St36 to attenuate depression-like and anxiety-like behaviors related to chronic stress.
Materials and Methods
Animals
The Georgetown University Animal Care and Use Committee approved both animal experiments in accordance with the standards set forth by the National Institutes of Health guidelines.
Experiment 1
Forty-two adult male, 11- to 12-week-old, Sprague-Dawley rats weighing 350–420 g were randomly assigned to 4 groups in this study: group 1 (n = 7), the control group, received no treatments; group 2 (n = 7) was assigned as the stress-only group that received the cold stress protocol; and groups 3 and 4 (n = 14 each) were both exposed to the cold stress protocol and then received either sham-EA or EA at St36, 30 minutes after the stress protocol. All animals had indwelling jugular vein catheters when obtained from Harlan Laboratories, Inc and were housed 1 per cage in a controlled environment at a constant temperature (23°C). Upon arrival and on day 7 of the experiment, catheter care was carried out injecting 0.1 mL of lock solution (5-mL 100% sterile glycerin [Fisher Scientific] and 5-mL 1000-U/mL heparin sodium) using a sterile 1-mL syringe with a blunt tip 23-gauge needle (Small Parts, Inc). All animals were maintained in a 12-hour light, 12-hour dark cycle with free access to water and regular rat chow.
Treatments
Before initiating the study the animals in all 4 groups (control, stress, sham-EA, and EA St36) were acclimated for 4 days by study personnel who handled them twice a day for 3 minutes. After the 3 minutes of handling, all the animals in the 4 groups were placed in a cotton sock headfirst, with their hind paws and tails exposed. The cotton sock was used in the study to briefly hold the animal in place only while inserting and taping the acupuncture needles for the treatment groups. Upon insertion of needles, the animals were then taken out of the sock and placed in single cages while receiving the acupuncture or sham acupuncture treatments. On day 5, the experimental treatments were initiated.
Stress treatments
Animals were placed a 1-cm deep ice/water mixture (0°C–3°C) for 1 hour daily for 14 days, as previously described (13), after which the rats were returned to their home cages, and their temperatures were allowed to normalize at room temperature (RT) for 30 minutes before initiating the sham-EA or EA St36 treatments.
EA treatments
To closely resemble the clinical setting, where patients receive acupuncture treatments for chronic stress after the stress has begun, EA at sham or St36 was initiated 30 minutes after daily cold stress exposure.
Sham-EA
After the animals had acclimated to RT for 30 minutes, they were placed in the sock with their hind paws and tails exposed, sterile 34-gauge (0.22 mm), 1-inche (25 mm) acupuncture needles (Millennia) were quickly inserted into nonacupuncture point on the back, 2 cm lateral to the tail region, bilaterally, and the animals were placed, unrestrained, in their individual cages. The needles were then attached to an acupuncture machine (model AWQ-104L, purchased from UPC Medical Supplies) via electrodes and were stimulated for 20 minutes at a frequency of 10 Hz with 2 mA output, as previously prescribed (9, 14, 15).
The rats in the EA St36 group received the same treatment as above, except that the needles were placed in the St36 point, located bilaterally on the hind leg of the rat lateral to the tibia, and just below the anterior tubercle of the tibia (9). Stomach 36 is a classic point used often in Chinese medicine for treating many disorders in humans, and has been reported to treat acute and chronic stress responses in the rat (9, 16). This point is also easily accessed in the conscious animal, minimizing the need for anesthetizing or immobilizing the animal, thus providing a useful model in studying stress.
Ten of the 14 rats in both the sham-EA and the EA St36 groups continued to receive EA for the 14 days that they were exposed to the cold stress. However, after 10 days, 4 of the animals in each of the sham-EA and EA St36 groups stopped receiving the EA treatments, whereas exposure to cold stress was continued for the remaining 4 days, to determine whether the effects of the sham-EA and/or EA St36 were long lasting.
Experiment 2
Thirty-eight adult male, 11- to 12-week-old, Sprague-Dawley rats weighing 350–420 g were randomly assigned to 5 groups in this experiment. Group 1 (n = 6) (control+vehicle) were animals that received no treatments and no RU-486 but on day 10 of the experiment received sc vehicle injections, consisting of 100 μL of dimethylsulfoxide. Group 2 (n = 6) (control+RU) was assigned as a control group that also received no treatment exposure (see control animals in Figures 6 and 7 below) but starting on day 10 of the experiment received sc RU-486 (1-g RU-486 [Sigma-Aldrich] was dissolved in 20-mL dimethylsulfoxide). Each animal was injected with 100 μL (0.1 mL) totaling a dose of 5 mg/animal per day sc. Animals in groups 3 (n = 6) and 4 and 5 (n = 10/group) were exposed to cold stress as described in experiment 1 for 9 days and on day 10 also received RU-486, 5 mg/d sc, for the remaining 4 days. Group 3 continued to get cold stress only (stress group) for a total of 14 days, whereas groups 4 (sham-EA+RU) and 5 (EA St36+RU) animals were exposed to the sham-EA or EA St36 treatments described above, respectively, for 14 days.
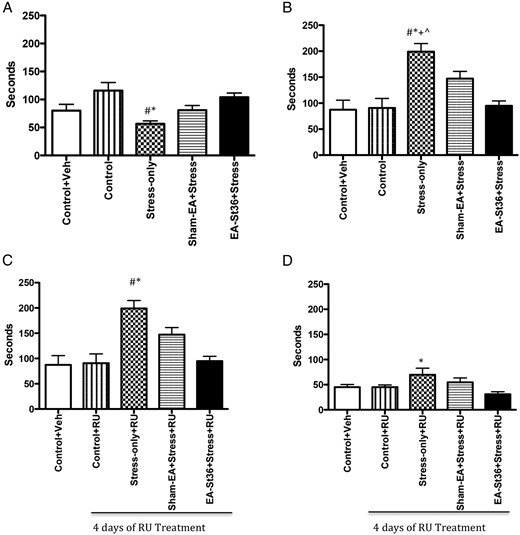
A, FST latency to immobility. Latency to immobility was significantly lower in the stress-only animals when compared with the 2 control groups and the EA St36 animals (*#, P < .05, respectively). B, FST duration of immobility. The duration of immobility in the stress-only animals was significantly higher (*#+^, P < .05) than those of all other animals, regardless of treatment. C, FST latency to immobility after treatment with RU-486. After 4 days of treatment with RU-486, the stress-only animals had significantly (*, P < .005) higher latency to immobility than the control animals or the EA St36 animals (#, P < .05). D, FST duration of immobility after treatment with RU-486. The duration of immobility was significantly higher (*, P < .05) in the stress-only animals when compared with control animals after treatment with RU-486
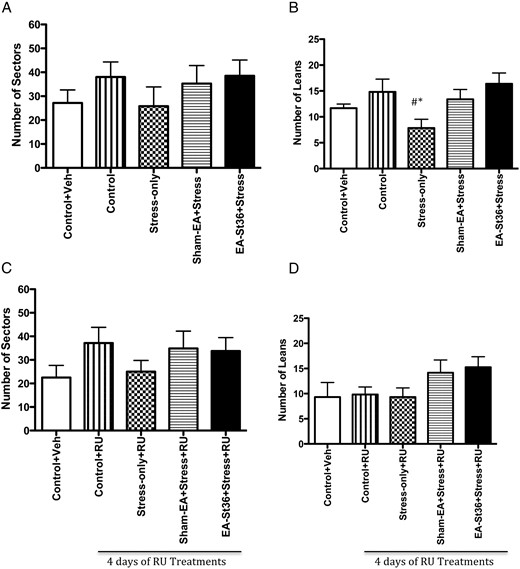
A, OFT sectors visited. Although the stress-only animals visited sectors the least demonstrating low locomotive activity, this was not significantly different than the other animals. B, OFT wall leans. The stress-only animals had significantly lower wall leans than the 2 control animal groups as well as the EA St36 animals (*#, P < .05). Conversely, this was not observed in the sham-EA animals. C and D, OFT sectors visited, wall leans after treatment with RU-486. Once the animals were treated with RU-486, no significant differences were noted in the OFT parameters, sectors visited, and wall leans
During this time, the animals were observed very carefully for any adverse effects from the combination of RU-486 and the cold stress. All animals survived the treatments.
Behavioral testing
Forced swim test (FST)
In order to assess depression-like behavioral responses, all of the 5 experimental groups were also tested in the FST, a well-established behavioral test for depression, immediately after the OFT (17–28). It has been examined as a suitable rodent model for measuring depression and further elucidating the effectiveness of antidepressants (29, 30). The FST, following the classic models, included a 5-minute period, where the animal was allowed to swim in a bucket filled with RT water. The animals could not reach the bottom of the bucket and were therefore forced to swim or float. Endpoints observed and measured in this paradigm included latency to immobility and duration of immobility.
Open field test (OFT)
All 5 groups of animals the experiments were exposed to the OFT, a commonly used measure of rodent anxiety (31–34). Rats were tested on day 10 of the stress/acupuncture protocol and then again after 4 days on RU-486. The test entailed allowing the animal to roam freely for 6 minutes, in a box divided with tape into sectors, each equal in dimensions and ambient light. Endpoints measured in this paradigm included sectors visited and wall leans, which refers to the number of times the animal demonstrates curiosity by leaning on the wall of the open field box with their forepaws while standing on their hind paws.
Blood sampling
Using the indwelling jugular vein catheters, 1–1.3 mL of blood were collected from each animal on day 1 (preexposure) of the experiment, and day 14 (final day of the study), using a 3-mL sterile syringe. To control for diurnal changes in HPA hormone levels, and provide for consistency across experiments, all blood samples were collected between 10 am and 12 pm. The collections occurred in the morning before killing the animals, 2 hours after the experimental exposures (stress and acupuncture treatments) were completed to allay acute stress measurements and to ensure uniform collection times. This is the same method used in our previous experiments, which also examined the chronic effect of our stress and acupuncture treatment exposures (9). Blood was placed in 1.5-mL EDTA tubes and centrifuged at 4°C at 6708g for 2 minutes. The plasma was stored at −80°C until analysis. Serum was collected for CORT measurement in a 1.5-mL low retention sterile tube, allowed to remain at RT for 15 minutes and centrifuged at RT for 5 minutes at 5000 revolutions per minute. The serum was stored at −80°C until analysis.
ELISA analysis of ACTH and CORT
The ELISA was performed per manufacturers' recommended protocols. ACTH ELISA kit (specificity of 100% for rat plasma ACTH with a typical sensitivity of 0.26 ng/mL, catalog S-1185; Bachem) was used to measure ACTH levels. The kit required an initial extraction step equilibrating a 200-mg C-18 sep-column with 1-mL 100% acetonitrile, followed by 1% trifluoroacetic acid (HPLC grade) 3 mL 3 times, followed by cold vacuum centrifugation. The collected residue was rehydrated with equal volume (150 μL) of assay buffer and used to analyze ACTH levels following the kit's protocol. CORT level determination was made using ELISA kit (with a specificity of 100% for rat CORT and typical sensitivity of 1.6 ng/mL, catalog DSL-10–81100; Diagnostic Systems Laboratories). The optical density from each assay was read on a standard ELISA plate reader set at 450 nm.
Immunohistochemistry
The paraventricular nucleus (PVN) of animals in experiment 1 was located from the dorsoventral coordinate of 8.0 mm from the bregma and mediolateral coordinate of 0.7 mm. Once identified, 5-μm sections at the level of the PVN were formalin fixed and paraffin embedded. Upon analysis, the sections were deparaffinized with xylenes and rehydrated via a graded alcohol series of 100%–70%. They were then rinsed in water and Tris-buffered saline with 0.05% Tween. The slides were immersed in 10mM citrate buffer (pH 6.0) with 0.05% Tween at 98°C for 20 minutes, then allowed to come to RT for 20 minutes; 3% hydrogen peroxidase and 10% normal goat serum were used for blocking. The slides were then incubated with a primary antibody, CRH (1:2000, ab8901; Abcam) for 1 hour at RT. Horseradish peroxidase-conjugated antirabbit secondary antibody (Dako Envision Plus) was applied for 30 minutes. The horseradish peroxidase was detected using 3,3′-Diaminobenzidine chromagen (Dako). Slides were counterstained with hematoxylin dilution for 2 minutes at RT, blued in 1% ammonium hydroxide for 1 minute at RT, dehydrated, and mounted with Acrymount. Negative controls were treated similarly except for exposure to the primary antibody. Slides were visualized using Cri Nuance FX microscope (Caliper Life Sciences). Images (×10) were saved using the Cri Nuance version 2.6.0 camera; CRH-positive staining was semiquantitatively analyzed by the Multidimensional Scaling Analytical Technologies Metamorph version 7.5.5.0.
Quantitative real-time RT-PCR
Quantitative real-time RT-PCR was used to determine PVN CRH mRNA expression in animals from experiment 2. Briefly, total RNA was isolated from resected brain paraventricular nuclei using the previously described phenol-chloroform extraction method (35). A total of 1 μg of RNA per sample was used for cDNA synthesis via the iScript cDNA synthesis kit (Bio-Rad). Real-time RT-PCR was done using ICycler iQ Detection System (Bio-Rad). The PCRs were carried out using TaqMan Universal PCR Master Mix and predesigned primers and fluorescein-labeled probes (Applied Biosystems). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), 5′-CCTTCATTGACCTCAACTAC-3′ and 5′-GGAAGGCCATGCCAGTGAGC-3′; CRH, 5′-CAA TCG AGC TGT CAA GAG AGC-3′ and 5′-GGA AGA AAT CCA AGG GCT GAG-3. The results were calculated by the comparative cycle time method using GAPDH for the brain as reference genes, per the Applied Biosystems ABI PRISM 7700 User Bulletin number 2.
Statistical analysis
GraphPad Prism version 4 (GraphPad Software) was used to analyze the data and presented as mean ± SEM. One-way ANOVA with Tukey multiple t test or Kruskal-Wallis post hoc test were used depending on sample size to compare between treatment groups. Significance was set at P < .05 for the indicated sample size per group.
Results
Experiment 1
Effects of EA St36 on stress-induced plasma ACTH and CORT in chronically stressed rats
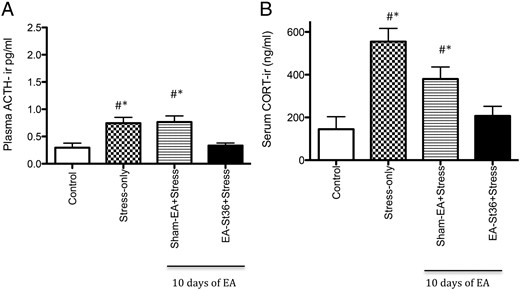
On day 1, before treatment of the experiment and day 7 (data not shown), plasma ACTH levels were not different between the 4 groups (range of 0.377–0.495 ng/mL). After 14 days of cold stress ACTH levels were significantly (P < .05) higher in the stress alone compared with the controls animals (Figure 1A). The stress effect on plasma ACTH in the stressed EA St36 animals was blocked and similar to levels in the nonstressed controls. In this animals, the plasma ACTH level was significantly lower than the stress-only and stress+sham-EA animals (P < .05) (Figure 1A).
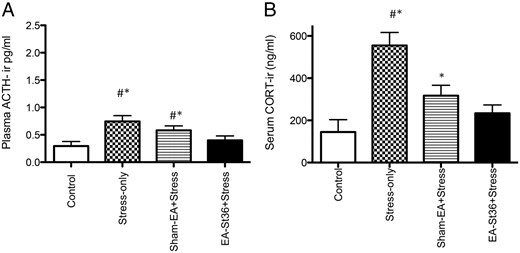
A, Plasma ACTH. Plasma ACTH was significantly higher in the stress alone and the sham-EA groups vs control animals (*, P < .05). Additionally, these groups of animals had significantly higher (#, P < .05) ACTH levels than the EA St36 rats. B, Serum CORT. Serum CORT levels were significantly higher in the stress alone and the sham-EA groups vs control animals (*, P < .05). Additionally, the stress-only group of animals had significantly higher (#, P < .05) CORT levels than the EA St36 rats, whereas there was no significant difference in levels between sham-EA and stress only
Mean serum CORT levels were not significantly among the 4 groups on pretreatment day 1 (data not shown). After 14 days of cold stress CORT levels (Figure 1B) were significantly elevated in the stress-only and stress+sham-EA animals compared with the unstressed-control group (P < .05), whereas there was no significant difference noted in the stressed animals that received EA St36 when compared with those from controls (P > .05). These effects were similar to our previous findings using pretreatment with EA St36 (9). Interestingly, there was no significant difference noted in CORT levels between the stress+sham-EA animals and the EA St36 group, and although the EA St 36 animals' CORT levels approached control animal levels, those of the stress+sham-EA animals did not and were indeed significantly higher as noted above.
Long-term effects of EA St36 on circulating ACTH and CORT
To determine whether the effects of EA St36 are long lasting, all EA treatments were stopped 4 days before the end of the experiment while the cold painful stress continued. ACTH levels in the stress-only and sham-EA+stress groups were significantly elevated compared with the unstressed control group (P < .05) (Figure 2A). Plasma ACTH in the EA St36 animals was still not different from the unstressed controls (Figure 2A), despite the discontinuation of the acupuncture treatment 4 days before the end of the experiment.
Longitudinal ACTH and CORT levels
Longitudinal ACTH and CORT level graphs demonstrate that, even when acupuncture was withdrawn while the cold stress treatments were continued for 4 days, both ACTH and CORT levels remained significantly higher in the stress-only and sham-EA animals in contrast to the EA-St 36 animals and the control animals (*#, P < .05, respectively).
We found a similar pattern in the CORT levels of the EA groups after stopping the acupuncture treatments but continuing the cold stress. This effect mirrored what we observed with ACTH, in that CORT levels in EA St36 animals were not different from unstressed control animal levels, whereas stress-only and stress+sham-EA levels were significantly elevated (P < .005) (Figure 2B). Thus, there is a long-term effect on ACTH and CORT that may be specific to EA St36, when compared with the sham-EA points.
Effects of EA St36 on stress-induced PVN CRH
CRH-immunoreactivity (ir) was significantly (P < .05) higher in the PVN of stress-only animals compared with control and both EA groups (Figure 3A). This was consistent with our previous findings (9). Figure 3B illustrates the average number of CRH-positive cells from each of the 4 groups. Chronic cold stress significantly increased CRH protein expression, whereas EA St36 prevented the increase in CRH. Although circulating CORT was elevated in the sham-EA animals, CRH-ir was not significantly higher than observed in control and EA animals.
A and B, PVN CRH-ir
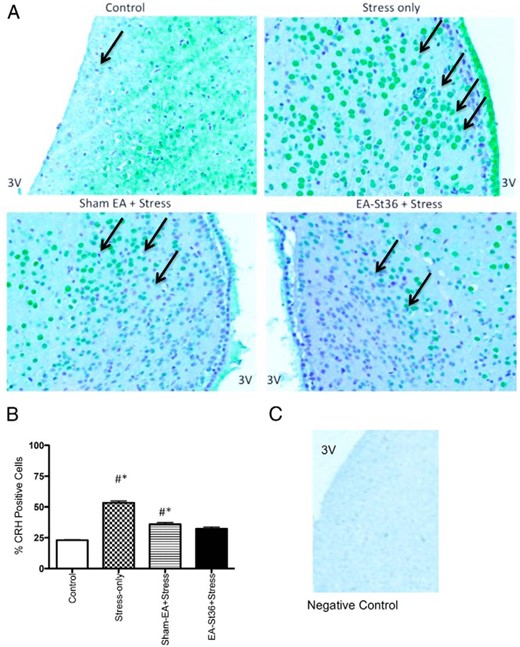
The percent of PVN CRH-positive cells in the stress-only animals was significantly higher than those of control and EA St36 animals (*#, P < .05, respectively). C, Technical negative control− PVN tissue was incubated with the antibody diluent, without the primary CRH antibody.
Experiment 2
When CORT activity was suppressed by using CORT receptor blocker, RU-486, plasma ACTH levels were similar to the patterns observed without GR blockade (Figure 1A). However, blocking GRs resulted in an exaggerated elevation in ACTH levels (mean of 1.98 ng/mL) in the stress-only+RU animals, compared with mean ACTH levels (0.837 ng/mL) of stress-only animals in experiment 1. The plasma ACTH levels of the vehicle only and RU-486-only groups were significantly lower than the stress and sham-EA groups' plasma levels (P < .0001) (Figure 4A). Treatment with EA St36 prevented the stress-induced elevations in ACTH, irrespective of the RU-486. In contrast, ACTH in sham-EA+RU animals remained significantly higher than controls+RU.
A, Plasma ACTH after treatment with RU-486. Plasma ACTH levels of the second experiment showing that after 4 days of RU-486 treatments, the stress-only animals and the sham-EA rats had significantly higher levels of ACTH than control+vehicle and control+RU animals (+*, P < .001, respectively). In contrast to the EA St36 animals, the sham-EA and stress-only animals demonstrated significantly (#, P < .001) higher levels of circulating ACTH. B, Serum CORT after treatment with RU-486. Serum CORT levels of the second experiment showing that after 4 days of RU-486 treatments, the stress-only animals had significantly lower levels of CORT than control+vehicle and control+RU animals (*+, P < .05, respectively). In contrast, circulating CORT was similar in the EA St36 animals, compared with the 2 control groups: control+vehicle and control+RU groups
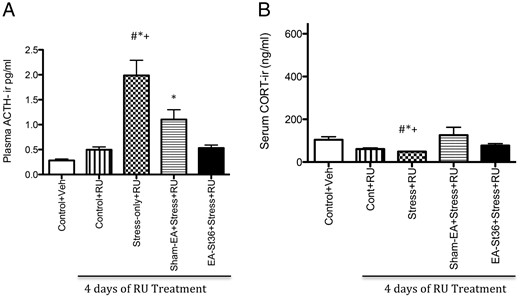
Blocking the GR with RU-486, during the final 4 days of stress exposure in experiment 2, abolished the increase in CORT observed in all of the chronically stressed rats (Figure 4B) and Eshkevari et al (9). In fact, as seen in Figure 4B, there were no significant differences from group controls in any of the serum CORT levels (despite the low levels in the stress-only RU animals), which is consistent with GR type I blockade (36).
PVN CRH in the presence of RU-486
Blocking the GR type I receptors using RU-486 prevented the elevation in CRH expression in the PVN of stress-only and sham-EA+stress animals previously noted in Figure 3, A and B. There was, in fact, no significant change in CRH protein expression; RU-486 had abolished the significant increase in CRH activity that was observed in experiment 1 in the stress-only animals (Figure 5).
PVN CRH expression after treatment with RU-486
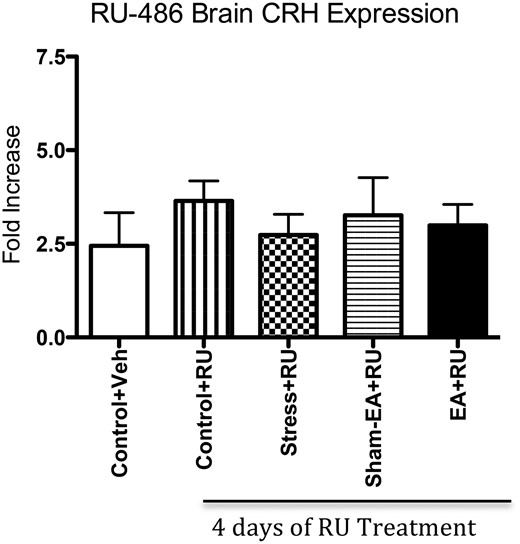
Examining CRH mRNA (normalized by GAPDH) after 4 days of RU-486 treatments revealed no significant variances among groups.
The behavioral effects of EA St 36 in chronically stressed rats with and without RU-486
There were several parameters measured in the FST. The first was the time at which the animal gave up and became immobile-measured as latency to immobility; the second parameter measured was the duration of the immobility when it did occur, denoting giving up and hopelessness in the animals. After chronic painful exposure to cold stress for 10 days, the animals in the stress-only group (Figure 6A) had significantly shorter latency time compared with both control and EA St36 animals (P < .05): they gave up quickly. In the second FST round of experiments, we examined duration of immobility (Figure 6B). Similar to the latency test, we found that those that were in the stress-only group had a significantly higher time, in which they had given up and were immobile as compared with the control and EA St36 animals (P < .05).
However, once the animals were treated with 4 consecutive days of RU while continuing with the stress and sham-EA or EA St36 protocols, the stress+RU group of animals had the highest latency to immobility rate. The control animals took significantly less time to become immobile compared with the stress+RU animals (P < .005). Interestingly, the EA animals behaved similarly to the control in that they too demonstrated significantly lower latency to immobility than the stress animals on day 10 (P < .05) (Figure 6C). Once we blocked the CORT receptor with RU (Figure 6D), we found that the overall duration of immobility in all animals was reduced, but that the stress+RU animals continued to have significantly longer duration of immobility when compared with the control and EA St36 animals (P < .05).
In order to examine the effects of acupuncture on anxiety related to chronic painful stress, we exposed the animals to an OFT and measured the number of sectors visited, wall leans, and general locomotion. We found that before the RU treatments, after 10 days of chronic cold stress the animals had no significant difference in the number of sectors visited (Figure 7A). However, the stress animals had significantly lower number of wall leans than the control and EA St36 animals, and this effect was not observed in the sham-EA point (Figure 7B). Once CORT blockade was implemented, the effects dissipated and all animals had similar number of sectors visited and wall leans, with no significant variations noted among the groups (Figure 7, C and D). The chronic stress animals in all 3 stress groups had similar sectors visited compared with the 2 control groups (Figure 7, C and D).
Discussion
In our previous work (9), we demonstrated that pretreatment with TCM EA at St36 prevented the stress-induced increases in HPA axis activity (9). We recognize that although some patients may use acupuncture as part of their preventative healthcare regimen, the current trend in its use is as an adjunct, or complementary to other modalities in treating specific diseases and health issues (8). Indeed, acupuncture, as with many other complementary and alternative interventions, is usually sought when patients experience a disruption in their health and/or wellbeing, in particular when traditional allopathic medicine alone has not provided adequate relief. To that end, the current study assessed the efficacy of acupuncture days after exposing the animals to the painful cold-stress protocol. We chose St36 because it is a potent acupuncture point, and because it has also been mapped from the human St36 to the rat model for experimental purposes and has been transposed from human leg to the rat hind-paw in several acupoint system models (37, 38).
We found that EA at St36, minutes after exposure to painful cold stress, was as effective in preventing elevations in HPA axis hormone as pretreatment with EA St36 (10). Furthermore, this suppression of stress-induced HPA axis may be specific to EA St36 acupuncture, because the animals that received sham-EA point stimulation displayed elevated ACTH and CORT. The clinical manifestations of acupuncture use in stress have been recently investigated, with some studies suggesting its effectiveness at managing stress-related symptoms in a variety of settings (10, 39–41). However, the mechanism of action has remained ambiguous, especially in settings where it is used after exposure to stress. The present study supports our previous work reporting that EA St36 acts by suppressing the HPA during chronic stress, and expands on that information by determining that EA St36 can work after initiation of chronic stress and that the effects are long lasting. The described effect of EA St36 on the HPA may, in turn, positively affect behavior and overall health. Indeed, our preliminary behavioral studies indicate that EA St36 may prevent depression, as observed in the FST. This is the first report linking the effects of EA St36 on the HPA to depressive and anxious behavior.
Of great importance to clinical therapy is that the effects of EA St36 are relatively long lasting, in that 4 days after EA treatment was withdrawn but cold stress continued, the HPA axis hormones remained at control levels in the EA St36 animals. This finding is also novel and provides a framework for potential clinical use for acupuncture. Current standard clinical therapy based on TCM theory occurs about 2 times a week for the first couple of weeks of treatment, followed by weekly treatments for a length of time depending on the disease or symptom being managed (42). The notion that the effects from acupuncture can last for at least 4 days in our stress model is congruent with clinical practice and can provide insight as to optimal time between treatments.
To further understand the effect of EA St36 on the HPA during chronic stress, we examined protein expression of CRH in the PVN. Our findings in the central nervous system mirrored the results obtained in the circulation of ACTH and CORT peripherally, with EA St36 preventing the stress-induced increase in CRH. This confirms our previous report (9) and extends it by demonstrating that the positive effect can be gained when EA is started while stress is ongoing. These findings provide further support for EA St36 in the treatment of stress-related mental disorders, such as posttraumatic stress disorder, as well as numerous depressive-type disorders. Indeed, it has been found that patients who suffer from major and atypical depression demonstrate diathetic CRH pathology (43–45). Given our observations of the effects of EA on stress induced CRH activity, we postulate that EA can potentially provide a minimally invasive, cost-effective option for management of these intractable disorders.
The present study also evaluated the effects on HPA feedback regulation and behavior during chronic stress by comparing EA St36 vs GR blockade (using RU-486) on the HPA. RU-486 was effective in suppressing CORT (Figure 4B) and PVN CRH (Figure 5). There was also an elevation in ACTH (Figure 4A) from the loss of CORT feedback, which has been observed by others who have studied the effects of RU-486 on the HPA and ACTH in various chronic stress models; the effect is believed to be due to a further dampening of the negative feedback system (46, 47). Interestingly, there was paradoxical and significant suppression of CRH in the stress-only+RU animals, which might be explained by an uncovering of tonic CRH suppression by another system. Other investigators have also found that pretreatment with RU-486 reduces CORT levels and minimizes variations among groups in several stress models (48, 49). One of the proposed mechanisms for this lowering of CORT with RU-486 treatments is that the drug has a direct effect at decreasing rat adrenal 3β-hydroxysteroid dehydrogenase/isomerase, 21-hydroxylase, and 11-hydroxylase production, which are all involved in glucocorticoid synthesis. Thus, by inhibiting these enzymes, CORT synthesis itself is affected, and a reduction in serum levels is observed (50). Due to this phenomenon, the animals were carefully observed during the cold-stress protocol to ensure that they could survive the stress. Another explanation for this may be one proposed by Dallman's group that during chronic stress CORT levels are sustained via a positive feed-forward loop (51). However, once the stress episode is over, systemic CORT and ACTH may be abnormally low (51). This, together with the RU-486 action on CORT, provides possible explanations for our results, consistent with other models of chronic stressed animals with CORT receptor blockade (46). Comparing the results of EA St36 and RU-486 on the HPA in our chronic cold stress model suggests that although they both impact the HPA, the EA St36 may have less side effects, because it appears to prevent the stress-induced elevations in ACTH and CORT, whereas RU-486 suppresses CORT but elevates ACTH. It also suggests that EA St36 has a primary central effect because the entire pathway is controlled centrally.
The use of GR blockers in stress-related disorders, such as depression has been suggested, however side effects from these treatments can be undesirable (52). To that end, we examined the role of EA St36 in the behavioral implications of chronic painful cold stress. We found that EA St36 was more effective when compared with the sham-EA in reducing depression-like behavior as tested by the FST paradigm. Specifically, latency to immobility and duration of immobility were significantly less in the EA St36 group when compared with the stress-only group. Blocking CORT receptors had minimal effect on this psychologically protecting effect in the duration of immobility test, but not in the latency to immobility where RU blockade mimicked plasma ACTH levels, again confirming the findings above. Further, anxiety-related behavior was examined using the OFT. We examined the number of sectors visited and vertical activity and found that pre-RU EA at St36 provided protection from anxiety, but once the CORT receptor blockade was implemented the effects disappeared, perhaps signifying the role of circulating CORT on chronic stress-related anxiety. These behavioral observations parallel our physiological markers, further suggesting that EA St36 helps mitigate the effects of chronic stress in our rodent model. This finding is congruent with findings in the literature regarding the importance of CORT in depression in both human and animal models (53, 54).
Several hypotheses exist as to the underlying mechanism of action of acupuncture, and the stimulation of acupoints in the body. Numerous studies have found that human and rat acupuncture points are associated with areas that are rich in nerve endings (38, 55, 56). Li et al (38) demonstrated that acupuncture points in rats were associated with higher density of nerve endings in skin and muscles where the points are located. It is further postulated that these fibers include A, and C afferent fibers of the pain pathways and play a role in the peripheral inhibition of pain by acupuncture (57, 58). Given that our model of stress is a painful cold stress, it is possible that the EA at St36 was indeed affecting peripheral pain pathways, allaying pain and stress responses. Additionally, central mechanisms for the modulation of pain pathways have also been elucidated in recent human and animal studies examining the various areas of the brain involved in these pathways, such as the periaqueductal gray, locus coeruleus, and projections from these areas to the PVN, and the PVN itself (59–61). This would explain our central response findings with the HPA, which is closely tied to the pain pathways in the aforementioned areas of the brain. Indeed, it is well established that CRH expressing neurons cross talk with neurons of the modulating pain pathways (62, 63). We therefore propose a potential mechanism that in our model of chronic painful cold stress-induced increase in stress hormones was allayed by the role acupuncture can play in pain pathways closely related to the HPA axis. Our current findings offer a novel approach, and by directly affecting the HPA axis without altering feedback systems, TCM EA may provide utility in management of clinical chronic stress-related disorders, with minimal side effects.
Conclusion
In summary, the present work reports that application of EA St36 after initiation of chronic stress prevents the stress-induced increases in HPA hormones, and has potentially long lasting effects. Furthermore, this action may be specific to EA St36 vs the sham points used, as sham-EA does not prevent the rise in stress hormones as effectively. EA St36. Behavioral testing indicates that during chronic stress, when EA St36 prevents the increases in HPA, it is associated with “normal” behavior, compared with more depressive and anxious behavior observed in stress-only animals (Figures 6, A and B, and 7B). Studies using glucocorticoid blockade (with RU-486) indicate that EA St36 probably does not act directly through the CORT/ACTH feedback system, but probably acts through central regulation of CRH. In all, the study strongly supports EA St36 as a therapeutic modality for treating chronic stress states.
Acknowledgments
This work was supported by an American Association of Nurse Anesthetists doctoral fellowship award (L.E.).
Disclosure Summary: The authors have nothing to disclose.
For News & Views see page 3483
Abbreviations
- CORT
corticosterone
- EA
electroacupuncture
- FST
forced swim test
- GAPDH
Glyceraldehyde 3-phosphate dehydrogenase
- GR
glucocorticoid receptor
- HPA
hypothalamus-pituitary-adrenal axis
- ir
immunoreactivity
- OFT
open field test
- PVN
paraventricular nucleus
- RT
room temperature
- SNS
sympathetic nervous system
- St36
stomach meridian point 36
- TCM
traditional Chinese medicine.
References

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}