




Abstract
Chromosome ‘painting’ refers to the hybridization of fluorescently labeled chromosome-specific, composite probe pools to cytological preparations. Chromosome painting allows the visualization of individual chromosomes in metaphase or interphase cells and the identification of both numerical and structural chromosomal aberrations in human pathology with high sensitivity and specificity. In addition to human chromosome-specific probe pools, painting probes have become available for an increasing range of different species. They can be applied to cross-species comparisons as well as to the study of chromosomal rearrangements in animal models of human diseases. The simultaneous hybridization of multiple chromosome painting probes, each tagged with a specific fluorochrome or fluorochrome combination, has resulted in the differential color display of human (and mouse) chromosomes, i.e. color karyotyping. In this review, we will summarize recent developments of multicolor chromosome painting, describe applications in basic chromosome research and cytogenetic diagnostics, and discuss limitations and future directions.
Introduction
The painting of a picture, similarly to the development of a scientific discipline, is an evolutionary process that often progresses, at times regresses, and frequently witnesses periods without much improvement or advance. However, there are times in which a reflection on the accomplished and as yet unaccomplished is appropriate. With the recent reports on the use of fluorescence in situ hybridization (FISH) to colorize entire genomes and distinguish all chromosomes with a specific hue, the painting of chromosomes has now reached such a point (1,2). For the first time, chromosome painting can be used to analyze the entire genome, allowing one to screen for chromosomal aberrations. Until now, cytogenetic screening tests for numerical and structural aberrations were restricted to conventional chromosome banding analyses.
Almost a decade ago, chromosome painting was developed independently by research teams at Lawrence Livermore National Laboratories (3) and at Yale University (4,5). Both groups had taken advantage of the availability of cloned DNA libraries that were derived from flow-sorted human chromosomes (6–9). The problem of chromosome specificity, which is incurred by the presence of ubiquitously distributed repetitive sequence motifs, was overcome by suppression hybridization. Suppression hybridization refers to the blocking of labeled, repetitive DNA with an excess of unlabeled, whole genomic DNA, or DNA fractions enriched in repetitive sequences, such as Cot-1 DNA. The cytogenetic community quickly realized the potential of chromosome painting to tackle tantalizing diagnostic problems, such as the identification of marker chromosomes and the reconstruction of complex chromosomal aberrations in tumor metaphases. Chromosome painting, however, has also become a versatile tool in basic research disciplines ranging from radiation biology, to evolutionary cytogenetics, and research dealing with aspects of the nuclear structure. Table 1 summarizes the broad spectrum of the above applications along with pertinent references (the list of references, due to the exponential growth of publications, remains rudimentary and refers to early applications of chromosome painting techniques).
Range of applications of chromosome painting
Chromosome painting probes have been improved rapidly and modified in several aspects. The first generation of probes, based on chromosome-specific phage libraries, were rather cumbersome to use, due to low insert-to-vector ratios which frequently resulted in a relatively high background staining. Some of these limitations were overcome with the availability of plasmid libraries where an improved insert-to-vector ratio and easier probe generation enhanced the painting quality considerably (8). However, in some instances, subregions of chromosomes notoriously were stained more weakly than others (e.g. the tip of chromosome 1p), which made the interpretations of hybridization results more difficult. More recently, two additional protocols for the generation of chromosome painting probes have become available.
(i) Based on chromosome flow sorting and subsequent PCR amplification using degenerate primers, high quality probes are accessible revealing a considerably improved signal-to-noise ratio, along with an improved staining homogeneity (34).
(ii) The same applies to probes generated by micro-manipulated dissection of normal metaphase chromosomes, again followed by sequence-independent DNA amplification (35–40).
Both approaches can be extended to produce probes for reverse chromosome painting, where aberrant chromosomes were either flow sorted or microdissected and subsequently used as painting probes on normal metaphase chromosomes in order to establish the origin of chromosomal material in marker chromosomes (41–43). Microdissection probes come with the distinct advantage that, in addition to whole chromosome painting probes, region-specific probes for chromosomal arms or chromosomal bands can be generated (44,45). Flow-sorted chromosome painting probes, however, have the advantage that the target number for subsequent DNA amplification can be expanded easily, thus ensuring a high complexity of the painting probes. Chromosome painting probes are now also available for an ever increasing number of species, most notably for the mouse (46–50) and the rat (51,52), allowing the expansion of chromosome painting analyses to animal models of human diseases (53). Lastly, the widespread use of chromosome painting in non-specialized laboratories has also become possible due to improved microscope hardware (microscopes and optical filters), the use of sensitive digital imaging devices (CCD cameras, confocal laser scanning microscopes) and an increasing number of suitable DNA haptenization and fluorescent labeling systems. Furthermore, all human chromosome libraries are now available, labeled and unlabeled, through commercial sources. Applied to the field of molecular cytogenetics, these developments form the basis for the recent achievement of the simultaneous color differentiation of all human and murine chromosomes (1,2,53,54).
Multicolor Chromosome Painting
One of the most attractive features of FISH is the possibility to distinguish, in a single experiment, multiple chromosomes or chromosomal targets simultaneously. The possibility of increasing the number of discernible targets has, of course, spurred the fantasy, and the goal to color-karyotype all human chromosomes has been a long perceived one. Ground work for multicolor FISH was laid out by research conducted in The Netherlands. Using three fluorochromes, 7-amino-4-methylcoumarin-3-acetic acid (AMCA), fluorescein isothiocyanate (FITC) and rhodamine, Nederlof et al. could show in 1989 that the simultaneous visualization of differentially labeled FISH probes is indeed possible (55). Shortly thereafter, an important technical twist was introduced—the same group could demonstrate that combinatorially labeled DNA targets could be discerned as well (56). Here, probes were labeled with either pure fluorochromes or with fluorochrome combinations, thus increasing the number of discernible targets beyond the number of fluorochromes. Experiments were also performed by not only employing fluorochrome combinations but also ratios of fluorochromes in which the percentage of a specific fluorochrome defines the probe origin (57). Using ratio labeling and visualization with triple bandpass optical filters, Dauwerse et al. could show in 1992, that up to 12 chromosomes could be visualized simultaneously (58). In 1993, Wiegant demonstrated that chromosome painting probes, directly conjugated to fluorochromes, can be used as well (59). The possibility of aiding the color differentiation with sensitive digital imaging devices enabled the extension of the spectrum of suitable dyes towards the near infrared. Here, up to seven different targets could be distinguished (60). However, finally in 1996, two teams demonstrated significant improvement in multicolor FISH by reporting the visualization of all 24 human chromosomes in specific colors (1,2).
Two alternative approaches were used to reach the goal of color karyotyping chromosomes: (i) fluorochrome-specific optical filters, termed m-FISH; and (ii) interferometer-based spectral imaging (introduced as spectral karyotyping or SKY). The filter-based approach was developed by Speicher and colleagues and employs the sequential image acquisition with five different fluorochrome-specific optical filters. Narrow band pass filters were designed to allow maximum distinction of fluorescent dyes. After a computer-based image shift correction, the image analysis software then calculates a chromosome segmentation mask based on the 4′,6-diamidino-2-phenylindole (DAPI) image. In this pre-defined area, the intensities for all subsequently acquired images will be recorded. Based on the labeling schemes, the program then decides whether a single pixel contains signals from one single fluorescent dye or from two or more fluorochromes. This information forms the basis for chromosome identification and color assignment (1).
Schröck et al. have utilized a different, novel concept for the imaging of multicolor experiments (2). Their image analysis is based on spectral imaging, which refers to a combination of spectroscopy, CCD imaging and conventional microscopy (61,62). Spectral imaging uses a Sagnac interferometer to generate a fluorochrome-specific optical path difference that, in turn, can be Fourier transformed to provide spectral information. Combined with sensitive imaging, utilizing a CCD camera, the fluorescence emission spectrum can be recovered simultaneously at all image points. Consequently, only one exposure of the fluorescent specimen is required for the differential color display of all chromosomes. Dedicated software then classifies the image, i.e. identifies pixels with identical spectra. Such pixels will be assigned the same classification color (which is obviously a pseudocolor solely chosen for maximal color separation of the chromosomes). Based on this classification, the actual karyotyping procedure is performed. An example is provided in Figure 1.
Applications of Multicolor Chromosome Painting
Clinical cytogenetics
The screening for chromosomal abnormalities in the pre- and postnatal laboratory is based on chromosome banding analyses. Today, known chromosomal disorders account for ∼5% of all children with birth defects or six in 1000 newborns (63). Growth retardation, a pattern of dysmorphic signs, malformations and mental retardation as well as multiple miscarriages and infertility frequently are caused by chromosome aberrations and are indications for chromosome analysis.
The major challenge in clinical cytogenetics is posed by the appearance of marker and ring chromosomes which cannot be identified by chromosome banding techniques alone (64,65). The mere presence of these marker chromosomes can be described by conventional banding analyses; yet, the chromosomal origin remains elusive. The characterization of these structures is important with respect to possible phenotypic consequences. Small marker chromosomes originating from chromosome 14 were reported to have nearly no phenotypic effect, but larger markers containing material from chromosome 15 indicate a high risk for malformation and mental retardation (66).
![SKY analysis of a patient with Wolf-Hirschhorn syndrome. (A) G-banded metaphase. The telomeric region of one chromosome 4 (arrow) shows a slightly abnormal banding pattern. (B) The same metaphase as shown in (A) after SKY analysis. Chromosome classification revealed an unbalanced translocation between chromosomes 4 and 8 [46,XY,der(4)t(4,8)].](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/hmg/7/10/10.1093/hmg/7.10.1619/2/m_7-10-1619-fig002.jpeg?Expires=1716388951&Signature=d~4dK6GjFy3fePB7~7~BgwxKuiUam5kwWeOyzrUVHEB8BqD4KBXlMAsd16AdHCTAm0osR-9fRv2sAO3FiPIuxdqSNwmJp21wisokbkwGAbnNJuZlpxYbhON4dkJOjiTXDNr1YbKXnOeUtX4Pc7An~Y-BpV0r6BjIJu9NRRteK16v3uoE1Ce8s9QtxDM34jk~Ete80Eg4txEylmjKG2e-S5SVhowWSfhPV4FzHgmNid4jeNp5bCdeiK0T6df2xseXaumIuq4WVfpwrW0gD5TkgDQzGq2dWwhLhldpn4EJwXniKZn5RA2UUq67dSPUisbz2MxuBOOUDABDeyqEGiE78w__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
SKY analysis of a patient with Wolf-Hirschhorn syndrome. (A) G-banded metaphase. The telomeric region of one chromosome 4 (arrow) shows a slightly abnormal banding pattern. (B) The same metaphase as shown in (A) after SKY analysis. Chromosome classification revealed an unbalanced translocation between chromosomes 4 and 8 [46,XY,der(4)t(4,8)].
FISH techniques have been developed and applied to identify the origin of the markers and other structural chromosomal aberrations (15,67,68). The use of chromosome painting probes in one, two or three color FISH experiments has significantly improved the definitive diagnosis of chromosomal aberrations. Many case reports underline the importance of this strategy for clinical cytogenetics (for review see ref. 69). This procedure is straightforward when conventional cytogenetic analyses suggest the involvement of specific chromosomes. Without any suspected involvement of specific chromosomal material, however, chromosome painting becomes a greater challenge because it requires the iterative hybridization of multiple chromosome painting probes. Therefore, chromosome painting is not always an option due to time constraints and limited specimen availability.
Additional strategies have been developed recently to accompany chromosome painting. Microdissection and cloning or PCR amplification of DNA sequences, followed by reverse painting, permit the detailed analysis of apparently aberrant chromosomes (70–72). These techniques have been transferred to advanced clinical laboratories in order to facilitate marker chromosome identification. However, these techniques lack the possibility of screening the whole genome.
Color karyotyping techniques hold the potential to facilitate the identification of chromosomal aberration in the pre- and postnatal cytogenetics laboratory without compromising one of the important features of cytogenetic diagnosis, i.e. the analysis of the entire genome. The power of color karyotyping techniques has been demonstrated in an extended study of clinical samples with suspected or unidentified, constitutional chromosomal abnormalities. Marker chromosomes, cryptic translocations and complex rearrangements could be characterized readily (2,73). In some patients afflicted with mental retardation and physical disabilities, spectral karyotyping unambiguously refined the cytogenetic diagnosis and identified specific chromosomal aberrations after repeated G-banding analyses had revealed a normal karyotype (74).
The limitations of chromosome painting probes, hybridized either singly or in a multicolor experiment, are obvious. The sensitivity of detecting interchromosomal structural aberrations, such as translocations, is much higher than for intrachromosomal aberrations: inversions or deletions are obviously harder to track because the color of the aberrant chromosomes remains unchanged. The combination of conventional banding analysis and color karyotyping helps to fill this methodological gap. An example of a comprehensive (in 1980) analysis of the same metaphase cell is presented in Figure 1. Conventional karyotyping performed in 1980 revealed a normal 46,XY karyotype. However, the phenotype of the patient was indicative of a Wolf-Hirschhorn syndrome. Subsequent high-resolution G-banding demonstrated an unusual banding pattern in chromosomal band 4p16.3. Spectral karyotyping eventually revealed a der(4)t(4,8). Approximately 90% of the patients afflicted with Wolf-Hirschhorn syndrome show a small deletion in this chromosomal band. Translocations have been described in ∼ 10% of the cases (75). Further methodological improvements also suggest that single-locus probe pools for frequently deleted chromosomal regions and for all human telomeres can be combined and microdeletion probe sets developed to identify submicroscopic microdeletions.
In summary, a genome-wide screening for chromosomal aberrations has become possible with the recent developments of color karyotyping. SKY is especially useful for detecting small translocations which are cytogenetically similar in appearance and to classify marker chromosomes and complex chromosomal aberrations. Finally, full automation of karyotype analysis in the clinical cytogenetic laboratory is within reach.
Tumor cytogenetics
The lower the quality of metaphase chromosomes and the higher the number of chromosomal aberrations, the more useful chromosome painting approaches are in helping to elucidate the pattern of chromosomal rearrangements. Clearly, one of the prime applications of m-FISH and SKY remains in tumor cytogenetics. In tumor metaphases with highly rearranged chromosomes, karyotype interpretation is often arduous because the shuffling of chromatin produces a banding pattern that obscures the original band sequence. Making matters worse, the mitotic index of tumor cell cultures frequently is very low, and clonal heterogeneity adds yet another level of complexity. The results of this enigma are a disturbingly high number of so-called derivative and marker chromosomes which are a mere reflection of the limitation of conventional banding approaches in the analyses of tumor metaphase chromosomes (76). Furthermore, in particular in solid tumor cytogenetics, cytogenetic features such as double minute chromosomes or homogeneously staining regions, which are chromosomal manifestations of oncogene amplification, are impossible to characterize using banding analyses alone. Chromosome painting alleviates the problem to a certain extent. The interpretation of complex karyotypes is facilitated further if all chromosomes are visualized at once in different colors rather than in subsequent individual single color chromosome painting experiments (Fig. 2) (77–79). In a recently conducted study, Veldman et al. could show that in 15 cases of hematological malignancies, the karyotype interpretation could be improved, completed or refined in all examples (80). The reasons for this improvement are manifold: (i) color karyotyping permits the identification of subtle telomeric translocations that exchanged chromatin of similar banding pattern, therefore escaping the description by banding techniques; (ii) small chromosomal markers, barely the size of a chromosomal band, could be identified; (iii) chromosomes thought to be normal were indeed identified as aberrant; and (iv) SKY allowed for the refinement of chromosomal breakpoint mapping, which is of tremendous importance in hematological malignancies because recurring chromosomal breakpoints often indicate the location of genes whose translocation results in overexpression or the generation of fusion proteins. It is likely to predict that the continued application of color karyotyping, again in conjunction with chromosome banding analyses, will result in the identification of hitherto unknown chromosomal aberrations in human cancers (81). Conceivably, a growing number of entry points for gene identification strategies and improved genetic markers for diagnosis, differential diagnosis, prognosis and therapy planning will result.
Comparative cytogenetics
The introduction of chromosome painting to the field of comparative cytogenetics (18) has added significantly to the understanding of chromosome changes that occurred during the evolution of species. Chromosome painting can be used to identify homologous chromosome segments in different species and to map probes of different complexities and chromosome rearrangements in a single experiment (22). In recent years, the complete karyotypes of various mammals including primates, carnivores and artiodaclys have been analyzed by chromosome painting (for a recent review see ref. 82). Most of the work used single chromosome painting probes hybridized to metaphases simultaneously analyzed by conventional G- and R-banding (19–21), replication banding (83) or DAPI banding (84–87).
A prerequisite for a rapid and profound analysis of chromosome changes which happened over evolutionary time is to cover the karyotype of the given species as completely as possible with molecular probes preferentially in a single experiment. The first mammalian karyotype, which was painted entirely with all chromosome-specific painting probes in a single hybridization experiment, was the Indian muntjac (2n = 6/7) (84). This deer species has the lowest chromosome number known in mammals. Painting probes for all chromosomes from the Indian muntjac were established by fluorescence-activated flow sorting, and the two autosome pairs and the X chromosomes were labeled with two haptens and a combination of them to achieve a three color pattern over the entire karyotype (Fig. 3A). Painting these probes to Chinese muntjac metaphase chromosomes, another closely related and morphologically similar deer species, but with a 2n = 46 karyotype, identified very simple changes in chromosome morphology. Single Indian muntjac paints hybridized to several entire Chinese muntjac homologs. Only two chromosomes were painted by more than one probe, indicating reciprocal translocations (Fig. 3B). The three color painting (red, yellow, green) readily demonstrated how the 2n = 6/7 karyotype of the Indian muntjac can be derived from the Chinese muntjac-like 2n = 46 ancestral karyotype by various tandem and centromeric fusions. This has been verified by using reciprocal painting with probes derived from the Chinese muntjac and painted to the Indian muntjac homologs (85,86).
![Detection of an interstitial deletion and terminal translocation der(5)t(5;12) by SKY in bone marrow cells of a patient afflicted with acute myeloid leukemia. This aberration was described as a terminal deletion of 5q [del(5)(q31)] using G-banding analysis. Additional rearrangements were identified as der(7)t(1;7), der(7;12), der(11)t(1;11) and der(21;22) (76). The normal homologs of the involved chromosomes are indicated for comparison.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/hmg/7/10/10.1093/hmg/7.10.1619/2/m_7-10-1619-fig003.jpeg?Expires=1716388951&Signature=QptEkHftMQqP9OA7IC8OabS4F2QBGhuHCjGBBSHpuqok61LUwqfwjs1mzEPaSEfR8vgVha6zmOcsoMSTYA~RiMICc2LT9jER0yKdi9t3ZrxosLV-Q-SPDV7FhL1f6q0HO1jkVNNEWcPFhdDdJByW408Buh8ssi1x5~ERNzeL4bQqkjkhY0J4zkzO4A37xGPXEyLb~Lh8Y090iH9laTmE7qLEQgP1d7JVEZ8sA7WWTfisfsez6whnDW29DKXIUmEsmxsxTt1T4kWSlv2~AnCWdeoIP-8ae3BdkKC~vw50UV9w8zjsuX-nSsZY42DThLv5bbzTPQBhnRcAl1YmoHiR4Q__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Detection of an interstitial deletion and terminal translocation der(5)t(5;12) by SKY in bone marrow cells of a patient afflicted with acute myeloid leukemia. This aberration was described as a terminal deletion of 5q [del(5)(q31)] using G-banding analysis. Additional rearrangements were identified as der(7)t(1;7), der(7;12), der(11)t(1;11) and der(21;22) (76). The normal homologs of the involved chromosomes are indicated for comparison.
Karyotype analyses of various gibbon species were performed by assigning the hybridization patterns from all 24 human chromosome-specific paints (21,87–89) in a series of single color chromosome painting experiments. All gibbon species are characterized by a highly disrupted chromosomal synteny. The profoundly shuffled gibbon genome is therefore considerably different from other hominoid primates, where only few translocation events are observed. With the exception of the sex chromosomes, banding analysis hardly allows one to identify homologous human and gibbon chromosomal regions. Even between the various gibbon species only few chromosomes seem to be conserved (90). Classical G-banding, in combination with subsequent hybridization with the human painting probes elucidated the numerous translocations. However, as for the analysis of highly rearranged human tumor chromosomes, the assignment of single chromosome paints to rearranged chromosomes and to banded chromosomes is laborious and requires confirmation.
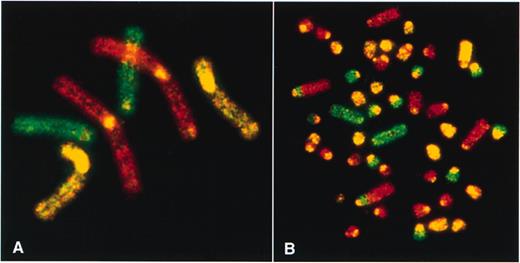
Conventional three color painting of metaphase spreads produced from Indian (A) and Chinese muntjacs (B) (kindly provided by Fengtang Yang, Cambridge, UK). Chromosome-specific painting probes were generated by flow sorting of a female Indian muntjac cell line, labeled by DOP-PCR and hybridized back onto Indian muntjac metaphase chromosomes (A: red signal, chromosome 1; green signal, chromosome 2; yellow signal, chromosomes 3; X and cross-hybridization signals of repetitive sequences to centromeres of most muntjac chromosomes). The 46 chromosomes of the Chinese muntjac (B) were painted entirely using the probes described above (with the exception of the Y chromosome). Therefore, the evolutionary changes between Chinese and Indian muntjacs could be described as the result of tandem and centromeric fusions and reciprocal translocations (84).
The entire karyotype of the Concolor gibbon (H.concolor) was analyzed recently by spectral karyotyping in a single SKY hybridization experiment (2). All of the ∼60 individual chromosome segments which were delineated with single human chromosome-specific painting probes were identified by spectral karyotyping. However, the results did more than simply confirm previous data. Some smaller chromosome rearrangements identified by spectral karyotyping have not been found in the previous single chromosome painting experiments, and the order of hybridization signals could be determined unambiguously. Spectral karyotyping also added information about chromosome polymorphisms in the karyotype of this particular individual, which would be difficult to achieve with single chromosome paints.
Conclusion
Chromosome painting has developed into an indispensable tool not only for chromosome analysis in basic research but also for diagnostic applications. In particular, the recent maturation of chromosome painting to color karyotyping can now be applied as hybridization-based karyotype analysis and as an initial screening test for chromosomal aberrations. It is likely that the majority of marker chromosomes in clinical and cancer cytogenetics will be readily identified. For diagnostic applications, color karyotyping will have to be combined with conventional chromosome banding analyses. This combination will greatly benefit from automated microscope hardware and software that permit the synergistic interpretation of results of the respective methodologies. Future work will obviously also be aimed at increasing the resolution of karyotype analysis by integrating chromosome arm and chromosome band-specific painting, and locus-specific, single-copy probe sets.
The application of color karyotyping to non-human species will offer rapid and easier karyotype analyses in species notoriously difficult to analyze, such as the mouse (53). This possibility will have a considerable impact on the understanding of basic mechanisms of chromosomal aberrations in animal models of human diseases and will shift a mainly descriptive technique into a functional one.
Acknowledgements
We are grateful to Fengtang Yang for providing Figure 3. The authors are indebted to Professor Malcolm A. Ferguson-Smith, Patricia O'Brien and Fengtang Yang (Cambridge, UK) for providing chromosome painting probes. The authors wish to thank Drs M. C. Phelan (Greenwood Genetic Center, Greenwood, SC) and J. C. Liang (Section of Cytogenetics, Division of Laboratory Medicine, The University of Texas M.D. Anderson Cancer Center, Houston, TX) for providing patient material. Joel Barnabas is gratefully acknowledged for critically editing the manuscript. This work was supported in part by Applied Spectral Imaging, Inc. (NIH CRADA, ASI 9512-8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}