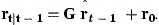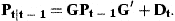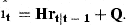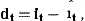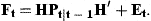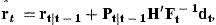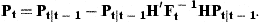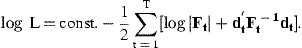Abstract
Catch-at-age analysis provides estimates of stock size at ages when the fish have reached catchable size. Survey indices contain information about relative cohort size at younger ages. The present analysis is concerned with survey indices of juveniles up to the youngest age where stock estimates, based on time-series analysis of catch-at-age data, are available. A stock estimate at that age from catch-at-age data is also included. A common model of the relationship between stock size and survey indices is combined with the model describing the decline of a stock by natural mortality. Random variations in natural mortality are defined separately from sampling variations and irregular catchability in the survey. The stock size and magnitudes of the random variations are estimated by the Kalman filter, which also provides predictions of future recruitment to the catchable stock. Analysis of observations of Icelandic cod reveals a large deviation from proportionality in the relationship between the index and the stock estimates in the youngest ages, but haddock data are compatible with proportionality. Variations in natural mortality during the second to fourth year of cod and the second to third year of haddock are not a major factor in variations of stock size.
1 Introduction
Survey indices provide information about relative stock size at the time of the survey. By combining this information with estimates of stock size from catch-at-age analysis, indices of younger fish can be used to predict recruitment to the catchable stock. Various statistical methods have been employed for this purpose (Rosenberg et al., 1992; Shepherd, 1997).
Myers and Cadigan (1993a, b) investigated survey indices of young fish from many stocks. Their analysis was mainly concerned with natural mortality and not with prediction of recruitment, and data or estimates from catch-at-age analysis were not included.
The present methodology is intended for stocks where annual survey indices can be obtained for a few years from each cohort before substantial exploitation begins. Catchability in surveys varies with age and size of the fish, particularly for the youngest ages. Before young fish enter the commercial fishery, their numbers decline due to natural mortality. The indices are subject to measurement errors and irregular variations in catchability. When a survey index is used to predict stock size at an older, catchable age, irregular variations in natural mortality between these ages contribute to the prediction error. With the time-series methodology, it is possible, at least theoretically, to estimate the magnitude of these variations separately from the irregular component of the survey indices.
We analyse in some detail abundance indices from a trawl survey on Icelandic cod in ages from 1 to 4 years from 1985 to 2002. For comparison with the results of Myers and Cadigan (1993a, b), we also include results from the analysis of haddock in ages from 1 to 3 years. The survey is carried out annually in March and was described by Pálsson et al. (1989). The cod data from the Marine Research Institute in Reykjavik (MRI, 2002, Table 3.1.10) are presented in Table 1. Estimates of the stock at 4 years of age, obtained by time-series analysis of catch-at-age data from the same report, are also included in Table 1. When the survey data become available, the last available catch-at-age data are from the previous year. The analysis is carried out in accordance with this, with catch-at-age results from 1985 to 2001. The main spawning period of Icelandic cod is in April. Notice that all fish hatched in year t are assigned age 0 during that year and 1 at the beginning of year t+1. Recruitment estimates provide additional information about the stock size at the youngest age in the catch-at-age analysis used in the management rules for this stock (Baldursson et al., 1996). Predictions made further ahead affect economic planning by fisheries and the government.
Survey indices of Icelandic cod for ages 1–4 and estimates of stock size at age 4 from catch-at-age analysis.
| Survey indices | |||||
|---|---|---|---|---|---|
| Age 1 | Age 2 | Age 3 | Age 4 | Stock (million fish) | |
| 1985 | 16.54 | 112.3 | 35.4 | 48.2 | 104 |
| 1986 | 15.10 | 61.0 | 95.7 | 22.5 | 108 |
| 1987 | 3.65 | 28.9 | 104.0 | 83.5 | 248 |
| 1988 | 3.45 | 7.5 | 72.7 | 104.9 | 206 |
| 1989 | 4.09 | 17.3 | 22.4 | 80.1 | 131 |
| 1990 | 5.57 | 12.1 | 26.7 | 14.3 | 68 |
| 1991 | 3.95 | 16.3 | 18.2 | 30.7 | 102 |
| 1992 | 0.72 | 17.5 | 33.8 | 19.1 | 75 |
| 1993 | 3.58 | 4.8 | 35.1 | 39.3 | 135 |
| 1994 | 13.76 | 16.0 | 8.9 | 27.1 | 98 |
| 1995 | 1.17 | 29.3 | 26.2 | 9.4 | 57 |
| 1996 | 3.63 | 5.4 | 41.9 | 28.3 | 126 |
| 1997 | 1.21 | 22.4 | 13.7 | 56.5 | 133 |
| 1998 | 7.98 | 5.5 | 29.8 | 16.0 | 73 |
| 1999 | 7.38 | 34.3 | 7.1 | 42.3 | 140 |
| 2000 | 18.81 | 28.5 | 55.6 | 7.2 | 46 |
| 2001 | 12.09 | 24.1 | 37.0 | 38.3 | 145 |
| 2002 | 0.91 | 38.7 | 41.2 | 40.3 | |
| Survey indices | |||||
|---|---|---|---|---|---|
| Age 1 | Age 2 | Age 3 | Age 4 | Stock (million fish) | |
| 1985 | 16.54 | 112.3 | 35.4 | 48.2 | 104 |
| 1986 | 15.10 | 61.0 | 95.7 | 22.5 | 108 |
| 1987 | 3.65 | 28.9 | 104.0 | 83.5 | 248 |
| 1988 | 3.45 | 7.5 | 72.7 | 104.9 | 206 |
| 1989 | 4.09 | 17.3 | 22.4 | 80.1 | 131 |
| 1990 | 5.57 | 12.1 | 26.7 | 14.3 | 68 |
| 1991 | 3.95 | 16.3 | 18.2 | 30.7 | 102 |
| 1992 | 0.72 | 17.5 | 33.8 | 19.1 | 75 |
| 1993 | 3.58 | 4.8 | 35.1 | 39.3 | 135 |
| 1994 | 13.76 | 16.0 | 8.9 | 27.1 | 98 |
| 1995 | 1.17 | 29.3 | 26.2 | 9.4 | 57 |
| 1996 | 3.63 | 5.4 | 41.9 | 28.3 | 126 |
| 1997 | 1.21 | 22.4 | 13.7 | 56.5 | 133 |
| 1998 | 7.98 | 5.5 | 29.8 | 16.0 | 73 |
| 1999 | 7.38 | 34.3 | 7.1 | 42.3 | 140 |
| 2000 | 18.81 | 28.5 | 55.6 | 7.2 | 46 |
| 2001 | 12.09 | 24.1 | 37.0 | 38.3 | 145 |
| 2002 | 0.91 | 38.7 | 41.2 | 40.3 | |
Survey indices of Icelandic cod for ages 1–4 and estimates of stock size at age 4 from catch-at-age analysis.
| Survey indices | |||||
|---|---|---|---|---|---|
| Age 1 | Age 2 | Age 3 | Age 4 | Stock (million fish) | |
| 1985 | 16.54 | 112.3 | 35.4 | 48.2 | 104 |
| 1986 | 15.10 | 61.0 | 95.7 | 22.5 | 108 |
| 1987 | 3.65 | 28.9 | 104.0 | 83.5 | 248 |
| 1988 | 3.45 | 7.5 | 72.7 | 104.9 | 206 |
| 1989 | 4.09 | 17.3 | 22.4 | 80.1 | 131 |
| 1990 | 5.57 | 12.1 | 26.7 | 14.3 | 68 |
| 1991 | 3.95 | 16.3 | 18.2 | 30.7 | 102 |
| 1992 | 0.72 | 17.5 | 33.8 | 19.1 | 75 |
| 1993 | 3.58 | 4.8 | 35.1 | 39.3 | 135 |
| 1994 | 13.76 | 16.0 | 8.9 | 27.1 | 98 |
| 1995 | 1.17 | 29.3 | 26.2 | 9.4 | 57 |
| 1996 | 3.63 | 5.4 | 41.9 | 28.3 | 126 |
| 1997 | 1.21 | 22.4 | 13.7 | 56.5 | 133 |
| 1998 | 7.98 | 5.5 | 29.8 | 16.0 | 73 |
| 1999 | 7.38 | 34.3 | 7.1 | 42.3 | 140 |
| 2000 | 18.81 | 28.5 | 55.6 | 7.2 | 46 |
| 2001 | 12.09 | 24.1 | 37.0 | 38.3 | 145 |
| 2002 | 0.91 | 38.7 | 41.2 | 40.3 | |
| Survey indices | |||||
|---|---|---|---|---|---|
| Age 1 | Age 2 | Age 3 | Age 4 | Stock (million fish) | |
| 1985 | 16.54 | 112.3 | 35.4 | 48.2 | 104 |
| 1986 | 15.10 | 61.0 | 95.7 | 22.5 | 108 |
| 1987 | 3.65 | 28.9 | 104.0 | 83.5 | 248 |
| 1988 | 3.45 | 7.5 | 72.7 | 104.9 | 206 |
| 1989 | 4.09 | 17.3 | 22.4 | 80.1 | 131 |
| 1990 | 5.57 | 12.1 | 26.7 | 14.3 | 68 |
| 1991 | 3.95 | 16.3 | 18.2 | 30.7 | 102 |
| 1992 | 0.72 | 17.5 | 33.8 | 19.1 | 75 |
| 1993 | 3.58 | 4.8 | 35.1 | 39.3 | 135 |
| 1994 | 13.76 | 16.0 | 8.9 | 27.1 | 98 |
| 1995 | 1.17 | 29.3 | 26.2 | 9.4 | 57 |
| 1996 | 3.63 | 5.4 | 41.9 | 28.3 | 126 |
| 1997 | 1.21 | 22.4 | 13.7 | 56.5 | 133 |
| 1998 | 7.98 | 5.5 | 29.8 | 16.0 | 73 |
| 1999 | 7.38 | 34.3 | 7.1 | 42.3 | 140 |
| 2000 | 18.81 | 28.5 | 55.6 | 7.2 | 46 |
| 2001 | 12.09 | 24.1 | 37.0 | 38.3 | 145 |
| 2002 | 0.91 | 38.7 | 41.2 | 40.3 | |
The relative magnitudes of the survey indices for cod at each age vary much more than the stock estimates. Higher irregular variations of the indices may have some part in this, but visual examination of Table 1 shows that it is mainly a systematic feature of the data; an index of a cohort that is large or small in any particular year in the indices, retains its relative position during the 4 years included here.
2 Model
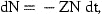
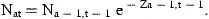
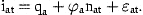
The actual age range will usually be from 0 or 1 to an age A, where estimates of stock size can be obtained from catch-at-age analysis. We cannot estimate absolute stock size in the ages below A from these data without some further information about the magnitude of catchability or mortality. No such knowledge will be assumed here.
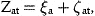
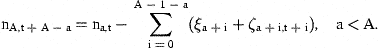
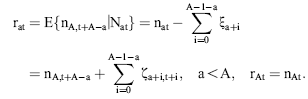
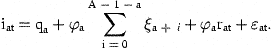
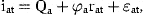
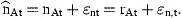
The errors, ɛn,t, are defined as normally distributed, independent of ɛat and serially uncorrelated.
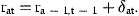
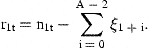
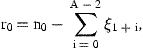
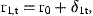
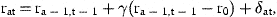
3 Estimation
The unobserved series of state variables, ra,t, are estimated from the observed indices and the catch-at-age estimates at age A by means of the Kalman filter. As the present models are linear in the state variables, no modification or extension of the Kalman filter is needed. This algorithm proceeds from initial values by predicting the state variables rat in the next period by their models with the expected value 0 inserted for the residuals δat. The predicted values of rat are used to predict the observed indices and 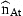 according to the models in Equations (7) and (8) and to calculate the prediction errors. These errors contain information about the estimates of state variables employed in the predictions. An overprediction of iat in Equation (7) indicates that rat was too large and should be adjusted. But the random element, represented by ɛat, also contributes to the prediction error of iat. If the variance of ɛat is large, compared with the uncertainty of the state variable, the adjustment should be small. The function of the Kalman filter is to determine the appropriate updating of the state variables in accordance with the prediction errors of the measurements and to calculate the covariance matrix of the updated estimates. A formal description of the application of the Kalman filter to the present model is given in Appendix 1.
according to the models in Equations (7) and (8) and to calculate the prediction errors. These errors contain information about the estimates of state variables employed in the predictions. An overprediction of iat in Equation (7) indicates that rat was too large and should be adjusted. But the random element, represented by ɛat, also contributes to the prediction error of iat. If the variance of ɛat is large, compared with the uncertainty of the state variable, the adjustment should be small. The function of the Kalman filter is to determine the appropriate updating of the state variables in accordance with the prediction errors of the measurements and to calculate the covariance matrix of the updated estimates. A formal description of the application of the Kalman filter to the present model is given in Appendix 1.
In order to apply the Kalman filter, we need initial values of the state variables in ages from 1 to A−1 years in the year before the first observations. All parameters in Equations (7), (8), (9), (12), and (13) must be known, including the full specification of the covariance matrices of δat and ɛat. In reality, these parameters are not known but can be estimated by the likelihood function of the prediction errors, i.e. we find the parameters by which the Kalman filter produces the best predictions of iat and 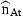 . Approximate standard deviations of estimated parameters are obtained from the Hessian matrix. The estimated covariance matrix of the state variables does not include uncertainty in parameter estimates and tends to underestimate the actual uncertainty, but not by an order of magnitude.
. Approximate standard deviations of estimated parameters are obtained from the Hessian matrix. The estimated covariance matrix of the state variables does not include uncertainty in parameter estimates and tends to underestimate the actual uncertainty, but not by an order of magnitude.
The direct estimation of the stock in Equation (8) is obtained by time-series analysis of the catch-at-age data (Gudmundsson, 1994). The best estimates of this stock at catchable ages are obtained from a joint analysis of these data and the survey indices. However, the uncertainty in specifying the correlation between ɛn,t from a joint analysis and the residuals ɛat in Equation (2) counteracts this advantage. In the present analysis, we use stock estimates obtained without using data other than the catch-at-age where we can safely assume that there is no correlation with ɛat. It is only in the last years that substantial differences may emerge between different estimation procedures.
Time-series estimation of nAt from catch-at-age data also provides estimates of the standard deviation of this estimate. But this estimation is rather inaccurate. It depends upon the method used for stabilizing variances and various somewhat arbitrary specifications of covariance matrices. Estimates of stocks and mortality rates in the last year are less accurate than in previous years, but the reverse applies to the assessment of the accuracy. In the present analysis, the standard deviation of ɛn,t in the last year, 2001, is obtained from the catch-at-age analysis. The standard deviation from 1985–1997 is estimated as a constant value σn, and the values from 1998–2000 as a linear interpolation between σn and the value in 2001.
The number of parameters is high. With four ages included there are two 4×4 covariance matrices, two parameters, Qa and ϕa, for each age in the measurement equations for the indices, and σn, r0 and γ. With T years of observations the size of T+A−1 cohorts must be determined, although only the A−1 initial values of rat enter as separate estimated parameters. The number of quantities to be determined is thus very high compared with the number of observations, but this is a common feature of cohort analyses of fish stocks.
It is necessary to introduce various simplifications in order to keep down the number of parameters and obtain well-determined estimates. The likelihood function is used to test the validity of parameter restrictions. The next section describes the specification and estimation with observations of Icelandic cod. Much of this will be common with data from other stocks, but not necessarily every detail.
The values of raT, obtained by the Kalman filter in the last year of observation, are our predictions of the recruitment to the stock at age A in the years from T to T+A−1, based on the whole data set. The Kalman filter estimates of rat in earlier years are based on the data up to and including year t. However, subsequent observations contain further information about these values. Estimates of rat, based on the whole data set, are obtained by a recursive procedure called “smoothing”.
Examination of residuals is an important aspect of applied time-series analysis. Observed residuals are the difference between the observed series and values predicted by the models. In the present analysis, the residuals are the prediction errors of iat and 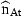 one year ahead. They are composed of the stochastic elements represented by ɛat and ɛnt in Equations (7) and (8) and also the errors produced by inaccurate estimation of the state variables rat. The residuals should be serially uncorrelated with all other residuals, but correlations of the prediction errors at different ages within the year are allowed in the model.
one year ahead. They are composed of the stochastic elements represented by ɛat and ɛnt in Equations (7) and (8) and also the errors produced by inaccurate estimation of the state variables rat. The residuals should be serially uncorrelated with all other residuals, but correlations of the prediction errors at different ages within the year are allowed in the model.
The prediction errors should have zero mean, but the variances differ in accordance with the different variances of the residuals in Equations (7), (8), (9), (12), and (13). The calculated residuals are therefore standardized by dividing by the calculated standard deviation of the respective residuals. Test statistics are calculated jointly for the standardized residuals from Equation (7). Two test statistics for normality are calculated, g1 for skewness and g2 for kurtosis. Both are approximately distributed as N(0;1) under the null hypothesis of normal distribution of the residuals. Two first-order serial correlation coefficients, rt and rc, are calculated. They represent the correlation between the residual at (a,t) with the residuals at (a,t−1) and (a−1,t−1), respectively. They are approximately normally distributed with zero mean and variances {A(T−1)}−1 and {(A−1)(T−1)}−1 under the null hypothesis of no serial correlation (Gudmundsson, 1986).
The Kalman filter, parameter estimation, and smoothing are described in textbooks on time-series analysis (e.g. by Harvey, 1989).
4 Examples
We describe here in some detail the parameter estimation and recruitment prediction of Icelandic cod. Parameter estimates for Icelandic haddock, obtained by similar methods, are also presented.
The survey indices of Icelandic cod in Table 1 are calculated in the same way at all ages. Most of the spawning takes place in the weeks immediately after the survey so the youngest age sampled in this survey is 1 year. There is a 0-group survey in August (Astthorsson et al., 1994). The 0-group index is more volatile than the trawl survey indices, and possible misspecification of the relationship with the stock values, including the stochastic aspect, could be damaging for the analysis. It is therefore not included in the present analysis, although the results convey some information about cohort strength.
The standard deviation of the stock estimation at 4 years of age in 2001 from the catch-at-age analysis is 0.17 and the present estimation of σn from 1985–1997 is 0.105 (Table 2). This estimate is too high according to the catch-at-age analysis, but as other results are not much affected if we use a lower value for this parameter, only results obtained with this value of σn will be presented. As 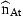 refers to the stock at the beginning of the year, the residuals ɛnt also include irregular mortality until the survey is carried out. But this is not a major factor because catches of 4-year-old cod are small during this period. The assumption of no serial correlation of the residuals in Equation (6) is incorrect for the last values, but they have bigger variances and thus exert less influence upon the results.
refers to the stock at the beginning of the year, the residuals ɛnt also include irregular mortality until the survey is carried out. But this is not a major factor because catches of 4-year-old cod are small during this period. The assumption of no serial correlation of the residuals in Equation (6) is incorrect for the last values, but they have bigger variances and thus exert less influence upon the results.
Parameter estimates and residual statistics. θ is estimated and γ is fixed in the first cod-column and in the haddock-column. In the second cod-column, θ is fixed and γ is estimated. Estimated standard deviations of the parameters are presented in parentheses below the respective parameter values.
| Cod | Cod | Haddock | |
|---|---|---|---|
| σɛ1 | 0.276 | 0.285 | 0.200 |
| (0.100) | (0.090) | (0.094) | |
| σɛ,2−A | 0.068 | 0.071 | 0.000 |
| (0.058) | (0.058) | (0.109) | |
| σκɛ | 0.159 | 0.158 | 0.214 |
| (0.037) | (0.038) | (0.098) | |
| σδ1 | 0.387 | 0.515 | 0.735 |
| (0.100) | (0.106) | (0.267) | |
| σδ,2−A | 0.069 | 0.072 | 0.082 |
| (0.026) | (0.030) | (0.085) | |
| σκδ | 0 | 0 | 0 |
| σn | 0.106 | 0.105 | 0.160 |
| (0.023) | (0.026) | (0.051) | |
| ϕ1 | 2.31 | 1.75 | 0.93 |
| (0.29) | (0.15) | (0.49) | |
| θ | −0.189 | 0 | 0.066 |
| (0.089) | (0.071) | ||
| γ | 0 | −0.091 | 0 |
| (0.035) | |||
| g1 | −0.61 | −0.61 | 0.08 |
| g2 | −1.48 | −1.49 | −1.29 |
| rt | 0.04 | 0.04 | −0.13 |
| rc | −0.17 | −0.15 | 0.10 |
| Cod | Cod | Haddock | |
|---|---|---|---|
| σɛ1 | 0.276 | 0.285 | 0.200 |
| (0.100) | (0.090) | (0.094) | |
| σɛ,2−A | 0.068 | 0.071 | 0.000 |
| (0.058) | (0.058) | (0.109) | |
| σκɛ | 0.159 | 0.158 | 0.214 |
| (0.037) | (0.038) | (0.098) | |
| σδ1 | 0.387 | 0.515 | 0.735 |
| (0.100) | (0.106) | (0.267) | |
| σδ,2−A | 0.069 | 0.072 | 0.082 |
| (0.026) | (0.030) | (0.085) | |
| σκδ | 0 | 0 | 0 |
| σn | 0.106 | 0.105 | 0.160 |
| (0.023) | (0.026) | (0.051) | |
| ϕ1 | 2.31 | 1.75 | 0.93 |
| (0.29) | (0.15) | (0.49) | |
| θ | −0.189 | 0 | 0.066 |
| (0.089) | (0.071) | ||
| γ | 0 | −0.091 | 0 |
| (0.035) | |||
| g1 | −0.61 | −0.61 | 0.08 |
| g2 | −1.48 | −1.49 | −1.29 |
| rt | 0.04 | 0.04 | −0.13 |
| rc | −0.17 | −0.15 | 0.10 |
σɛa: measurement errors and variations in catchability at age a; σκɛ: joint variations in catchability; σδa: irregular variations in natural mortality at age a; σκδ: joint irregular variations in natural mortality; σn: errors in stock estimates from catch-at-age analysis at 4 years' age; ϕ1: constant in a model of stock-dependent catchability; θ: coefficient of age in a model of stock-dependent catchability; γ: coefficient in age-dependent natural mortality; g1: test for skewness, N(0;1); g2: test for kurtosis, N(0;1); rt: test for first-order serial correlation in time, N(0;0.122) for cod, N(0;0.142) for haddock; rc: test for first-order serial correlation within cohorts, N(0;0.142) for cod, N(0;0.182) for haddock.
Parameter estimates and residual statistics. θ is estimated and γ is fixed in the first cod-column and in the haddock-column. In the second cod-column, θ is fixed and γ is estimated. Estimated standard deviations of the parameters are presented in parentheses below the respective parameter values.
| Cod | Cod | Haddock | |
|---|---|---|---|
| σɛ1 | 0.276 | 0.285 | 0.200 |
| (0.100) | (0.090) | (0.094) | |
| σɛ,2−A | 0.068 | 0.071 | 0.000 |
| (0.058) | (0.058) | (0.109) | |
| σκɛ | 0.159 | 0.158 | 0.214 |
| (0.037) | (0.038) | (0.098) | |
| σδ1 | 0.387 | 0.515 | 0.735 |
| (0.100) | (0.106) | (0.267) | |
| σδ,2−A | 0.069 | 0.072 | 0.082 |
| (0.026) | (0.030) | (0.085) | |
| σκδ | 0 | 0 | 0 |
| σn | 0.106 | 0.105 | 0.160 |
| (0.023) | (0.026) | (0.051) | |
| ϕ1 | 2.31 | 1.75 | 0.93 |
| (0.29) | (0.15) | (0.49) | |
| θ | −0.189 | 0 | 0.066 |
| (0.089) | (0.071) | ||
| γ | 0 | −0.091 | 0 |
| (0.035) | |||
| g1 | −0.61 | −0.61 | 0.08 |
| g2 | −1.48 | −1.49 | −1.29 |
| rt | 0.04 | 0.04 | −0.13 |
| rc | −0.17 | −0.15 | 0.10 |
| Cod | Cod | Haddock | |
|---|---|---|---|
| σɛ1 | 0.276 | 0.285 | 0.200 |
| (0.100) | (0.090) | (0.094) | |
| σɛ,2−A | 0.068 | 0.071 | 0.000 |
| (0.058) | (0.058) | (0.109) | |
| σκɛ | 0.159 | 0.158 | 0.214 |
| (0.037) | (0.038) | (0.098) | |
| σδ1 | 0.387 | 0.515 | 0.735 |
| (0.100) | (0.106) | (0.267) | |
| σδ,2−A | 0.069 | 0.072 | 0.082 |
| (0.026) | (0.030) | (0.085) | |
| σκδ | 0 | 0 | 0 |
| σn | 0.106 | 0.105 | 0.160 |
| (0.023) | (0.026) | (0.051) | |
| ϕ1 | 2.31 | 1.75 | 0.93 |
| (0.29) | (0.15) | (0.49) | |
| θ | −0.189 | 0 | 0.066 |
| (0.089) | (0.071) | ||
| γ | 0 | −0.091 | 0 |
| (0.035) | |||
| g1 | −0.61 | −0.61 | 0.08 |
| g2 | −1.48 | −1.49 | −1.29 |
| rt | 0.04 | 0.04 | −0.13 |
| rc | −0.17 | −0.15 | 0.10 |
σɛa: measurement errors and variations in catchability at age a; σκɛ: joint variations in catchability; σδa: irregular variations in natural mortality at age a; σκδ: joint irregular variations in natural mortality; σn: errors in stock estimates from catch-at-age analysis at 4 years' age; ϕ1: constant in a model of stock-dependent catchability; θ: coefficient of age in a model of stock-dependent catchability; γ: coefficient in age-dependent natural mortality; g1: test for skewness, N(0;1); g2: test for kurtosis, N(0;1); rt: test for first-order serial correlation in time, N(0;0.122) for cod, N(0;0.142) for haddock; rc: test for first-order serial correlation within cohorts, N(0;0.142) for cod, N(0;0.182) for haddock.
For Icelandic cod, we use A=4 years and must then specify and estimate the 4×4 covariance matrix of the residuals ɛat in Equation (2). These residuals represent measurement errors and variations in catchability. We know from analysis of catch-at-age data and survey indices that estimation of joint variations in catchability from ages 4 to 9 years yields large and highly significant values (Gudmundsson, 1998). The convenient prior assumption of independent residuals is therefore inappropriate.
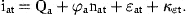
The standard deviation of ɛ1t, representing measurement errors and catchability variations in the survey for the 1-year-old fish, is much higher than for the older fish. The possibility that the large variations at 1 year of age are not highly correlated with variations in the older ages was investigated by excluding κat when a=1 and found to fit better. The results for cod presented in our tables were obtained with this option. The residual variations at older ages are highly correlated, indicating that variations in catchability are more important than sampling variations. The possibility of defining the covariance matrix with correlations declining with increasing age-difference was also investigated, but the present formulation was easier to estimate and fitted slightly better.
Free estimation of the ten parameters, σɛa, σκɛ, σδa, and σκδ produces rather badly determined values for most parameters. But the restriction to use the same parameter for the standard deviation of ɛat for ages 2–4 years and another for δat in these ages is acceptable (log L drops by 2.9).
According to the estimated values (Table 2), the variations of natural mortality, represented by σδ,2–4, are much lower than variations in stock size at age 1, represented by σδ1. The variations in cohort size at 4 years of age are therefore dominated by variations in spawning and natural mortality in the first year. There is no indication of correlation within each year between the residuals δat, associated with variations in natural mortality, regardless of the choice of specification.
We are estimating recruitment to A=4 years for cod in this study but there are also some recorded catches in younger ages. The effect of correcting for catches at 3 years was investigated but had negligible effect upon the goodness-of-fit and did not improve it. The correction was not applied in the results presented here. Unrecorded discards are equivalent to natural mortality in this analysis. Although we cannot determine relative variations in catchability in the younger ages without knowing the natural mortality rate, it is obvious from Table 1 that catchability at 1 year of age is much lower than in the higher ages.
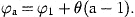
The smoothed estimates of rat, obtained with γ=0, are presented in Table 3, transformed from logarithmic values to stock size in numbers. The variations in these values within each cohort represent the estimated effects of the irregular variations in natural mortality, denoted by δat in Equation (9). The same estimates, obtained with stock-dependent natural mortality, are presented in Table 4. The variations within cohorts in this table are the estimated effects of stock-dependent and irregular variations in natural mortality.
Smoothed estimates of exp(rat) for Icelandic cod when variation with age of stock-dependent catchability is estimated.
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 201 | 225 | 101 | 117 |
| 1986 | 154 | 211 | 217 | 101 |
| 1987 | 77 | 152 | 221 | 217 |
| 1988 | 94 | 73 | 162 | 215 |
| 1989 | 92 | 94 | 77 | 156 |
| 1990 | 107 | 90 | 101 | 71 |
| 1991 | 104 | 106 | 84 | 106 |
| 1992 | 54 | 106 | 112 | 81 |
| 1993 | 101 | 55 | 107 | 117 |
| 1994 | 145 | 103 | 57 | 102 |
| 1995 | 64 | 142 | 105 | 57 |
| 1996 | 107 | 65 | 137 | 111 |
| 1997 | 59 | 111 | 67 | 139 |
| 1998 | 139 | 58 | 107 | 70 |
| 1999 | 131 | 143 | 50 | 121 |
| 2000 | 145 | 133 | 144 | 47 |
| 2001 | 158 | 137 | 132 | 139 |
| 2002 | 56 | 161 | 132 | 131 |
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 201 | 225 | 101 | 117 |
| 1986 | 154 | 211 | 217 | 101 |
| 1987 | 77 | 152 | 221 | 217 |
| 1988 | 94 | 73 | 162 | 215 |
| 1989 | 92 | 94 | 77 | 156 |
| 1990 | 107 | 90 | 101 | 71 |
| 1991 | 104 | 106 | 84 | 106 |
| 1992 | 54 | 106 | 112 | 81 |
| 1993 | 101 | 55 | 107 | 117 |
| 1994 | 145 | 103 | 57 | 102 |
| 1995 | 64 | 142 | 105 | 57 |
| 1996 | 107 | 65 | 137 | 111 |
| 1997 | 59 | 111 | 67 | 139 |
| 1998 | 139 | 58 | 107 | 70 |
| 1999 | 131 | 143 | 50 | 121 |
| 2000 | 145 | 133 | 144 | 47 |
| 2001 | 158 | 137 | 132 | 139 |
| 2002 | 56 | 161 | 132 | 131 |
Smoothed estimates of exp(rat) for Icelandic cod when variation with age of stock-dependent catchability is estimated.
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 201 | 225 | 101 | 117 |
| 1986 | 154 | 211 | 217 | 101 |
| 1987 | 77 | 152 | 221 | 217 |
| 1988 | 94 | 73 | 162 | 215 |
| 1989 | 92 | 94 | 77 | 156 |
| 1990 | 107 | 90 | 101 | 71 |
| 1991 | 104 | 106 | 84 | 106 |
| 1992 | 54 | 106 | 112 | 81 |
| 1993 | 101 | 55 | 107 | 117 |
| 1994 | 145 | 103 | 57 | 102 |
| 1995 | 64 | 142 | 105 | 57 |
| 1996 | 107 | 65 | 137 | 111 |
| 1997 | 59 | 111 | 67 | 139 |
| 1998 | 139 | 58 | 107 | 70 |
| 1999 | 131 | 143 | 50 | 121 |
| 2000 | 145 | 133 | 144 | 47 |
| 2001 | 158 | 137 | 132 | 139 |
| 2002 | 56 | 161 | 132 | 131 |
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 201 | 225 | 101 | 117 |
| 1986 | 154 | 211 | 217 | 101 |
| 1987 | 77 | 152 | 221 | 217 |
| 1988 | 94 | 73 | 162 | 215 |
| 1989 | 92 | 94 | 77 | 156 |
| 1990 | 107 | 90 | 101 | 71 |
| 1991 | 104 | 106 | 84 | 106 |
| 1992 | 54 | 106 | 112 | 81 |
| 1993 | 101 | 55 | 107 | 117 |
| 1994 | 145 | 103 | 57 | 102 |
| 1995 | 64 | 142 | 105 | 57 |
| 1996 | 107 | 65 | 137 | 111 |
| 1997 | 59 | 111 | 67 | 139 |
| 1998 | 139 | 58 | 107 | 70 |
| 1999 | 131 | 143 | 50 | 121 |
| 2000 | 145 | 133 | 144 | 47 |
| 2001 | 158 | 137 | 132 | 139 |
| 2002 | 56 | 161 | 132 | 131 |
Smoothed estimates of exp(rat) for Icelandic cod when variation with age of stock-dependent natural mortality is estimated.
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 254 | 266 | 101 | 117 |
| 1986 | 176 | 245 | 234 | 101 |
| 1987 | 69 | 165 | 238 | 217 |
| 1988 | 91 | 68 | 170 | 215 |
| 1989 | 87 | 93 | 74 | 156 |
| 1990 | 109 | 87 | 100 | 71 |
| 1991 | 105 | 107 | 82 | 106 |
| 1992 | 44 | 106 | 113 | 81 |
| 1993 | 101 | 48 | 108 | 117 |
| 1994 | 161 | 103 | 53 | 102 |
| 1995 | 55 | 151 | 105 | 57 |
| 1996 | 109 | 59 | 142 | 111 |
| 1997 | 49 | 112 | 64 | 139 |
| 1998 | 156 | 52 | 108 | 70 |
| 1999 | 143 | 154 | 47 | 122 |
| 2000 | 160 | 140 | 149 | 47 |
| 2001 | 183 | 146 | 135 | 138 |
| 2002 | 46 | 177 | 136 | 131 |
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 254 | 266 | 101 | 117 |
| 1986 | 176 | 245 | 234 | 101 |
| 1987 | 69 | 165 | 238 | 217 |
| 1988 | 91 | 68 | 170 | 215 |
| 1989 | 87 | 93 | 74 | 156 |
| 1990 | 109 | 87 | 100 | 71 |
| 1991 | 105 | 107 | 82 | 106 |
| 1992 | 44 | 106 | 113 | 81 |
| 1993 | 101 | 48 | 108 | 117 |
| 1994 | 161 | 103 | 53 | 102 |
| 1995 | 55 | 151 | 105 | 57 |
| 1996 | 109 | 59 | 142 | 111 |
| 1997 | 49 | 112 | 64 | 139 |
| 1998 | 156 | 52 | 108 | 70 |
| 1999 | 143 | 154 | 47 | 122 |
| 2000 | 160 | 140 | 149 | 47 |
| 2001 | 183 | 146 | 135 | 138 |
| 2002 | 46 | 177 | 136 | 131 |
Smoothed estimates of exp(rat) for Icelandic cod when variation with age of stock-dependent natural mortality is estimated.
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 254 | 266 | 101 | 117 |
| 1986 | 176 | 245 | 234 | 101 |
| 1987 | 69 | 165 | 238 | 217 |
| 1988 | 91 | 68 | 170 | 215 |
| 1989 | 87 | 93 | 74 | 156 |
| 1990 | 109 | 87 | 100 | 71 |
| 1991 | 105 | 107 | 82 | 106 |
| 1992 | 44 | 106 | 113 | 81 |
| 1993 | 101 | 48 | 108 | 117 |
| 1994 | 161 | 103 | 53 | 102 |
| 1995 | 55 | 151 | 105 | 57 |
| 1996 | 109 | 59 | 142 | 111 |
| 1997 | 49 | 112 | 64 | 139 |
| 1998 | 156 | 52 | 108 | 70 |
| 1999 | 143 | 154 | 47 | 122 |
| 2000 | 160 | 140 | 149 | 47 |
| 2001 | 183 | 146 | 135 | 138 |
| 2002 | 46 | 177 | 136 | 131 |
| Million fish | ||||
|---|---|---|---|---|
| Year | 1-yr olds | 2-yr olds | 3-yr olds | 4-yr olds |
| 1985 | 254 | 266 | 101 | 117 |
| 1986 | 176 | 245 | 234 | 101 |
| 1987 | 69 | 165 | 238 | 217 |
| 1988 | 91 | 68 | 170 | 215 |
| 1989 | 87 | 93 | 74 | 156 |
| 1990 | 109 | 87 | 100 | 71 |
| 1991 | 105 | 107 | 82 | 106 |
| 1992 | 44 | 106 | 113 | 81 |
| 1993 | 101 | 48 | 108 | 117 |
| 1994 | 161 | 103 | 53 | 102 |
| 1995 | 55 | 151 | 105 | 57 |
| 1996 | 109 | 59 | 142 | 111 |
| 1997 | 49 | 112 | 64 | 139 |
| 1998 | 156 | 52 | 108 | 70 |
| 1999 | 143 | 154 | 47 | 122 |
| 2000 | 160 | 140 | 149 | 47 |
| 2001 | 183 | 146 | 135 | 138 |
| 2002 | 46 | 177 | 136 | 131 |
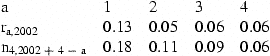
The last row of Table 3 shows the predictions of the stock size at 4 years of age in the years 2005, 2004, 2003, and 2002, obtained from the survey values of 2002. Table 5 shows similar predictions obtained with survey indices ending in 1998–2002 and the corresponding stock estimates ending in 1997–2001. The specifications were the same as in Table 3, but the parameters were reestimated for each data set.
Recruitment predictions of Icelandic cod at age 4 years.
| Million fish | |||||
|---|---|---|---|---|---|
| exp(r1T) | exp(r2T) | exp(r3T) | exp(r4T) | 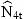 | |
| 1998 | 127 | 62 | 106 | 67 | 73 |
| 1999 | 126 | 143 | 55 | 111 | 140 |
| 2000 | 189 | 138 | 148 | 50 | 46 |
| 2001 | 151 | 143 | 135 | 141 | 145 |
| 2002 | 56 | 160 | 132 | 131 | |
| σ(pred) | 0.18 | 0.11 | 0.09 | 0.06 | |
| Million fish | |||||
|---|---|---|---|---|---|
| exp(r1T) | exp(r2T) | exp(r3T) | exp(r4T) | | |
| 1998 | 127 | 62 | 106 | 67 | 73 |
| 1999 | 126 | 143 | 55 | 111 | 140 |
| 2000 | 189 | 138 | 148 | 50 | 46 |
| 2001 | 151 | 143 | 135 | 141 | 145 |
| 2002 | 56 | 160 | 132 | 131 | |
| σ(pred) | 0.18 | 0.11 | 0.09 | 0.06 | |
The Kalman filter provides estimates of the standard deviation of raT but the error in the prediction of the logarithmic value of the stock at 4 years of age also includes the uncertainty caused by irregular natural mortality. This is included in σ(pred).
Recruitment predictions of Icelandic cod at age 4 years.
| Million fish | |||||
|---|---|---|---|---|---|
| exp(r1T) | exp(r2T) | exp(r3T) | exp(r4T) | | |
| 1998 | 127 | 62 | 106 | 67 | 73 |
| 1999 | 126 | 143 | 55 | 111 | 140 |
| 2000 | 189 | 138 | 148 | 50 | 46 |
| 2001 | 151 | 143 | 135 | 141 | 145 |
| 2002 | 56 | 160 | 132 | 131 | |
| σ(pred) | 0.18 | 0.11 | 0.09 | 0.06 | |
| Million fish | |||||
|---|---|---|---|---|---|
| exp(r1T) | exp(r2T) | exp(r3T) | exp(r4T) | | |
| 1998 | 127 | 62 | 106 | 67 | 73 |
| 1999 | 126 | 143 | 55 | 111 | 140 |
| 2000 | 189 | 138 | 148 | 50 | 46 |
| 2001 | 151 | 143 | 135 | 141 | 145 |
| 2002 | 56 | 160 | 132 | 131 | |
| σ(pred) | 0.18 | 0.11 | 0.09 | 0.06 | |
The Kalman filter provides estimates of the standard deviation of raT but the error in the prediction of the logarithmic value of the stock at 4 years of age also includes the uncertainty caused by irregular natural mortality. This is included in σ(pred).
For comparison with these results and the results from Myers and Cadigan (1993a, b) we also analysed haddock indices from the same survey. The time period was the same but the age range was from 1 to 3 years. The estimated standard deviation of the log-value of the stock at 3 years of age in 2002 from catch-at-age analysis was 0.23. Catchability of haddock at 1 year of age is much higher than for cod and the joint variations in catchability, represented by κɛt, were included at all ages. No evidence of stock-dependent catchability or natural mortality appears in the haddock results, i.e. ϕ1 is not significantly different from 1 and θ and γ are not significantly different from 0.
The test statistics show no evidence of non-normality or serial correlation in the analyses of these data sets.
5 Discussion
The data employed in the present work could also be used with other methods currently applied to predict recruitment. But unless random variations of natural mortality and joint catchability variations are negligible, these methods are not fully efficient because assumptions of independent residuals in their specifications are violated.
Our description and examples of this procedure were in terms of indices from a single survey. The basic theory is the same with indices from more than one survey for some or all of the ages, provided they are carried out at the same time. In view of the large correlations between the residuals associated with the present survey indices, the possibility of correlations between indices of different surveys should be investigated and taken into account. There are no methodological difficulties in carrying this out.
Joint analysis of indices from surveys, carried out at different times in the year, requires more extensive modification of the methodology. In order to keep variations in natural mortality separate from measurement errors and variations in catchability, the state variables rat should be updated at the time of each survey so we would no longer be working with time intervals of 1 year. However, the results of Myers and Cadigan (1993b) and the present paper indicate that irregular variations in natural mortality may often be so small that it is harmless to ignore them in recruitment predictions. It is not necessary that each survey includes all ages or that they all start in the same year.
Our results for cod differ in two respects from those of Myers and Cadigan (1993a, b). We find a small but significant irregular variation in natural mortality and a strong stock-dependence in catchability. In both cases, the difference rests upon the information in the series of stock estimates from the catch-at-age data, included in our analysis but not in the work of Myers and Cadigan. If we reduce the effect of this series by assigning a bigger variance (σn2) to the stock estimates, the estimated variance in natural mortality (σδ,2–42) becomes insignificant. Myers and Cadigan (1993a) assume that the estimated abundance in the survey is proportional to the true abundance, i.e. ϕ1=1. According to our results, this assumption is untenable for Icelandic cod. However, this conclusion also rests upon the information from the catch-at-age estimation. It was carried out with the traditional assumption of a constant natural mortality (0.2 year−1) in the catchable ages. If this assumption is wrong and the natural mortality is in fact strongly stock-dependent, then the stock estimates, based on the assumption of a constant natural mortality, are systematically wrong and the evidence for stock-dependent catchability could vanish. It is difficult to estimate the magnitude or the variations in natural mortality in ages with high fishing mortality. At the moment, we have no presentable evidence for or against the hypothesis of stock-dependent natural mortality of Icelandic cod in the catchable sizes and ages (>50 cm, ≥4 years).
For Icelandic haddock, we find no significant irregular variation in natural mortality. The precision of this estimation is lower for haddock than for cod because we have one age less in the survey data and less accurate catch-at-age estimates. It is therefore not possible to infer from the present analysis that the actual variations are in fact smaller for haddock than for cod. There is no indication of stock-dependent catchability or natural mortality for haddock. This is in agreement with the results from most of the haddock stocks investigated by Myers and Cadigan (1993a). In view of the difference between the results from Icelandic cod and haddock in these respects, it is worth noting that catchability in our survey is much higher for haddock than for cod. The stock in numbers of cod is bigger than the stock of haddock, but the survey catches five times more haddocks than cods.
Appendix 1
The Kalman filter algorithm
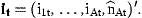
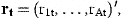
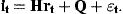
The coefficient matrix H is determined by Equations (7) and (8). The diagonal elements are ϕa in the first A rows. The Ath element in row A+1 is 1. All remaining elements are 0. The first A elements of the vector Q are Qa and the last is 0. The residuals have covariance matrix Et. The first A rows and columns are occupied by the covariance matrix of the residuals ɛat. The last diagonal element is the variance of ɛn,t and the off-diagonal elements in the last row and column are 0 if the stock estimates 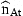 are independent of the survey indices.
are independent of the survey indices.
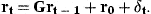
The coefficient matrix G has elements g21=…=gA,A−1=1 and the remaining elements are 0. The first element of the vector r0 is r0 and the remaining elements 0. The covariance matrix of δt is Dt.
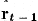 with covariance matrix Pt−1. We can then predict the next state variables by
with covariance matrix Pt−1. We can then predict the next state variables by