





Abstract
Zooplankton samples were collected by Hensen net (300 μm) vertical hauls from a depth of 50 m in 57 surveys along the Peruvian coast during winter and summer from 1961 to 2001. In general, the latitudinal distribution of total zooplankton volumes showed highest values within 60 nmi (111 km) of the coast. Predominantly during the 1960s, high zooplankton volumes were found between 4°S–6°S and 14°S–16°S, coinciding with narrow continental shelf areas. Strong declines of zooplankton volumes were observed in these regions in the 1990s. In the long term, zooplankton volumes off Peru were higher during “cold” decades, particularly in the 1960s, than during “warm” decades. This research supports the regime-shift previously observed in the Peruvian coastal upwelling, and is in agreement with global changes observed in lower and upper trophic levels elsewhere in the Pacific and North Atlantic basins.
Introduction
The Peruvian marine ecosystem is characterized by strong and persistent upwelling along the Peruvian coast, producing well-known biological productivity and supporting one of the largest fisheries in the world. Oceanographic and atmospheric conditions off Peru are characterized by several modes that vary on seasonal to interdecadal scales. One of the dominant modes is interannual variability due to the El Niño–Southern Oscillation cycle (ENSO), which affects ecosystem structure, reproductive strategies, and fisheries yields (Arntz and Valdivia, 1985; Bakun, 1987; Jarre-Teichmann, 1992). Interdecadal changes have also been observed and deduced from variability of sea surface temperature (SST), among other climatic variables, and linked to widely fluctuating catches of the Peruvian anchoveta (Engraulis ringens) and sardine (Sardinops sagax) (Chavez et al., 2003). A multidecadal trend of increasing wind velocity off Peru as a consequence of global warming has been proposed (Bakun, 1990). Three ENSO modes have been detected in the Pacific basin, long term, interdecadal, and interannual (Enfield and Mestas-Nuñez, 2000), and it is probable that many biological and physical time-series from the Pacific basin are related with these ENSO modes.
A decline in the California Current zooplankton (Roemmich and McGowan, 1995) and in the northern part of the Subarctic Pacific (Sugimoto and Tadokoro, 1998), from the start of the 1970s onward, has been suggested. Carrasco and Lozano (1989) also showed a decline in zooplankton volumes from the mid-1960s and especially from the mid-1970s to 1987 in the Peruvian upwelling ecosystem. Changes in physical forcing, e.g. winds, thermocline, heat flux, and SST, have been proposed (Clarke and Lebedev, 1999) to explain the decline in the Californian waters, and predation upon zooplankton of pelagic fishes, such as horse mackerel (Trachurus picturatus) and mackerel (Scomber japonicus), (Carrasco and Lozano, 1989) for the Peruvian waters. However, global changes associated with an interdecadal regime-shift in the dominant small pelagic fishes in the Pacific basin (Lluch-Belda et al., 1992; Schwartzlose et al., 1999; Hare and Mantua, 2000; Chavez et al., 2003) constitute the general framework in which we should try to understand trends in zooplankton volumes.
The current data set can be used to examine whether such changes persist off Peru. In Peruvian coastal waters, environmental factors and predation intensity may both play a crucial role in zooplankton fluctuations. Interdecadal variability of SST has been detected in this region (Montecinos et al., 2003) and the main fish predators of zooplankton have also shown interdecadal changes (Csirke et al., 1996; Schwartzlose et al., 1999). This paper is an extension of the proposal made by Carrasco and Lozano (1989) to synthesize the temporal and spatial trends in zooplankton biomass off Peru. We focus on regime-shifts of SST during the 1960–1970 and 1980–1990 decades, and interdecadal fluctuations of anchoveta biomass.
Data and methods
The total area covered during the study was 9.3×104 km2, from the coast to 556 km (300 nmi) offshore (Figure 1). Between 1964 and 2002, 130 seasonal surveys were carried out to assess anchoveta abundance and more than 10 000 zooplankton samples were collected by the Instituto del Mar del Peru (IMARPE). Zooplankton samples were taken with Hensen nets of 0.33-m2 mouth area and 300-μm mesh, by vertical hauls between 50 and 0 m. The samples were fixed in 2% formaldehyde buffered with borax. The zooplankton volume (m−3) was determined immediately after collection using the displacement method (Kramer et al., 1972). Ichthyoplankton and large coelenterata were separated from the samples before measurement. All the data come from the database of the Zooplankton Laboratory of IMARPE; a more detailed analysis, including species composition, has been initiated. As the sampling protocol has been consistent since 1964, we observed that similar proportions of day and night samples have been collected during each cruise. For spatial analysis, the Peruvian coast has been divided into three areas: A (03°30′S–5°59′S), B (06°00′S–13°59′S), and C (14°00′S–18°20′S) according to differences in oceanographic characteristics.
Study area along the Peruvian coast with a total of 6625 sampling points from summer and winter surveys from 1964 to 2002.
Monthly mean SST was measured in situ from 1950 to 1999 at five coastal stations along the Peruvian coast [Talara (04°34′S), Chicama (07°41′S), Chimbote (09°04′S), Callao (12°03′S), and Ilo (17°38′S)]. Data for the most recent years were not available from this source. We also show the mean annual series of SST from 1964 to 2001, from the IMARPE database. This mean annual SST was the result of averaging mean monthly SST of Paita (05°04′S), Chimbote, Callao, and Ilo coastal stations of IMARPE. The annual mean anchoveta biomass (Guevara-Carrasco, unpublished) comes from an updated version of the virtual population analysis (VPA) made by Csirke et al. (1996).
Statistical descriptors
The zooplankton data consist of 4 columns (seasons) and 39 rows (years). In this matrix, observations represent the mean value of each survey. The mean annual volume was obtained by averaging the rows (ZAM). Of the 39 years considered in the study, 31 have more than two surveys with complete spatial coverage, 7 years have one survey, and 1 year has no survey (1988). The surveys with incomplete spatial coverage (1979 and 1987) were discarded. These three annual mean values were replaced by a 5-year moving average of the annual series.
Time-series analysis
The survey frequency for zooplankton over the years and by season is shown in Figure 2. Monthly (PC1M) and annual (PC1A) SST data time-series were composed of the first principal component for five coastal stations. The PC1M and PC1A include most of the coastal variability during the 1950–1999 period for monthly and annual frequencies (600 and 50 observations, respectively). We analysed PC1A with a “low-pass filter” (moving average >7 years) in order to enhance “cold” and “warm” periods between 1964 and 1994.
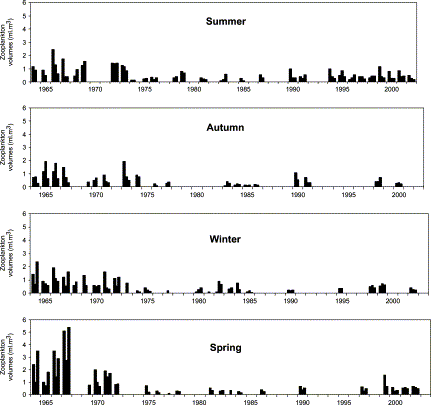
Zooplankton mean volumes collected during four seasons along the Peruvian coast. Each bar represents one cruise.
Spatial analysis
For spatial zooplankton variation, we used 3564 and 3065 sampling points collected during summer and winter, respectively, from 1964 to 2002. Data collected during spring and autumn had large gaps and were not considered. We explored decadal spatial variation using between 5 and 13 surveys per decade. The data were plotted and interpolated with the “closed neighbour” method using Surfer v.8.0 (Figure 3). An EOF analysis was performed but it did not increase the information. Data were averaged for seasonal comparisons among decades.
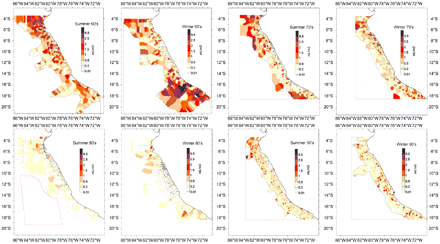
Spatial variability of zooplankton volumes, by decade and season, from 1962 to 2002. Contours were made by the closed neighbour method of interpolation. Sampling points are shown.
Results
The mean zooplankton volume ranged from 0.03 ml m−3 to 5.37 ml m−3. The seasonality of zooplankton volume did not show drastic changes among decades. Spring has been the season with the highest values among years in the 1960s and 1970s. A lesser contrast among seasonal volumes was observed during the 1980s and 1990s (Figure 4). The spatial distribution of zooplankton volumes showed the highest values from the coast to 60 nmi (111 km) offshore, between 04°S and 06°S and between 14°S and 16°S, coincident with narrow shelf areas and strong upwelling events (Zuta and Guillen, 1970). Important differences among areas (latitude) were detected. Although three areas had declining trends in zooplankton volumes (Figure 5), those of the northern (A) and southern (C) areas had more drastic declines between the 1960s and 1990s (65% and 70%, respectively) than the central area (B), which showed a reduction of only 55% in the same period.
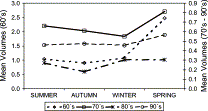
Mean seasonal zooplankton volumes for four decades for four seasons. Spring has always shown the highest values even during periods of less contrast during the 1980s and 1990s. Note that the scale of the ordinate in the 1960s differs from that of the 1970–1990s.
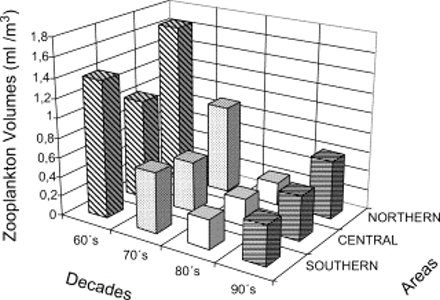
Mean spatial pattern of zooplankton volumes among decades indicates a dramatic decline in northern and southern areas.
The long-term trend of annual mean zooplankton volume, from 1964 to 2001, indicated three different periods of abundance: highest from 1964 to 1973, lowest from 1974 to 1989 (although the recovery seems to start in 1985), and an intermediate level from 1990 to the present (Figure 6). For comparison, the annual time-series of anchoveta mean biomass is shown. From 1964 to 1983 a significant match between two series can be observed with a high linear correlation (r2=0.58; p<0.01; ANOVA test); however from 1984 to 2001 there was no significant correlation.
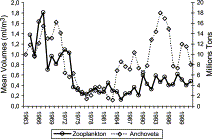
Mean annual zooplankton volumes and Peruvian anchoveta biomass from 1963 to 2001. Zooplankton values of 1979, 1988, and 1989 were interpolated with a 5-year moving average.
The trend of smoothed PC1A of SST showed alternating “cold” and “warm” periods (Figure 7). Two clear changes of phase (cold–warm–cold) have occurred in the environmental characteristics of the Peruvian upwelling ecosystem, as indicated by SST analysis. One change occurred in the mid-1970s and the other by mid-1980s. It is also possible to postulate a third change of phase at the start of the 1990s (cold–warm), but this is still uncertain because of recent alternations of cold years (1995–1996) and warm years (1997–1998). No significant correlation between mean zooplankton volumes and annual mean SST was detected. However, there was a coincidence between changes of phase of SST and changes in periods of abundance of zooplankton volumes and anchoveta biomass (Figures 6 and 7).
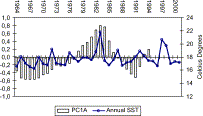
Trend of smoothed first principal component of sea surface temperature (SST) from five coastal stations from 1964 to 1994, to observe “cold” and “warm” regimes. Mean annual SST is also shown (compare with Figure 6).
Discussion
This paper is a contribution to the knowledge of long-term zooplankton dynamics in the Pacific basin. Trends observed in different components of the Peruvian ecosystem are in general agreement with changes observed over the entire basin (Chavez et al. 2003). Zooplankton represents one of the key elements of the marine ecosystem. This updated time-series of biomass in the Peruvian upwelling ecosystem has been made possible following active rescue of archival samples. Although “volume” (ml m−3) is not the best measure of biomass, it is the only one available at the moment in such long time-series. We recognize that the data are sparse in some periods. However, the quantity and quality of the data are probably the best available in the southeast Pacific, taking into account that all these samplings were part of surveys for the assessment of Peruvian anchoveta abundance and not specifically directed to the assessment of zooplankton.
It is clear that SST does not explain the changes observed in zooplankton volumes by itself, but can be a good indicator of what is happening with other oceanographic and atmospheric variables. For this reason, even when the correlation between biological variables and SST was not significant, its usefulness for interpreting observed changes is very important. According to an interannual analysis of ENSO indices (Trenberth and Stepaniak, 2001), the SST near the international date line has a different trend with respect to the SST along the coast of the Americas in the tropics. At the same scale, Montecinos et al. (2003) found a highly significant positive correlation between the SST of the American coast and that of the central Pacific. However, on an interdecadal scale the SST of the Peruvian coast and the PDO was highly significantly negatively correlated (−0.65, p<0.05); thus, according to these authors, the Peruvian system has cold and warm regimes, slightly out of phase with the total signal for the Pacific basin. The change of phase between these regimes (Figure 7) resulted in a notable agreement with those observed in biological variables (Figure 6).
The relationship between zooplankton and the anchoveta series is quite interesting. Even when a predator–prey relationship is known to exist, the ability of anchoveta to switch from phytoplankton to zooplankton under adverse environmental conditions (Rojas de Mendiola, 1989), and vice versa, precludes the establishment of an inverse correlation between zooplankton and anchoveta. In general, both series presented show three approximately congruent periods of different abundance: higher, lower, and intermediate. While it is clear that a good match exists between the two series from 1964 to 1983, they have some differences from 1984 to the present. Csirke et al. (1996) postulated three levels of carrying capacity for anchoveta or what could now be called “regimes”: the highest occurred in the 1960s until the collapse in 1972, the lower regime occurred from 1973 to 1983, and an intermediate level occurred from 1984 to 1994, now updated by this work to 2001 (Figure 6). During the recent period, anchoveta had 5 years (1992–1996) of high abundance that were similar to those of the 1960s, but after that it seems to have stayed at the “intermediate” level. Zooplankton also had three periods of different abundances, but the period of “intermediate” abundances started later (1990) than that of anchoveta (1984), and the zooplankton volumes have increased from the mid-1980s to the present at a lower rate than those of anchoveta.
At the moment, we do not have a sound explanation for these observations, but we should consider that the Peruvian anchoveta is only one species while the zooplankton is a complex of interacting species with a taxonomic composition that probably changes through time. Considering the type of net used to collect samples, the data of this study represent only changes of some in the taxonomic groups of the total zooplankton community. Different taxa can respond to environmental changes in different ways. For example, Nakata and Hidaka (2003) report that trends in biomass of large species of copepods respond better to global climate variables, while biomass of small species of copepods respond better to local climate variables.
Sánchez (2000) showed trends in total plankton volumes, collected with a phytoplankton net, in relation to distance from the coast from 1976 to 1999. Samples collected within the first 30 nmi of the coast corresponded mainly to phytoplankton, while samples taken beyond 30 nmi offshore contained more microzooplankton (S. Sánchez, pers. comm.). According to this study, plankton volumes decreased from 1976 to 1986 and increased from 1988 to 1999, both inside and outside 30 nmi offshore. This trend also showed two periods of different abundance, in similar ways to those of the anchoveta and zooplankton. If plankton volumes from the inshore represent a reasonable index of phytoplankton abundance, then the same observed trends in the series of three components could have important consequences for the understanding of the functioning of the Peruvian upwelling ecosystem.
Considering the spatial distribution of zooplankton over the decades studied, a strong decrease in zooplankton total volume in both northern and southern areas was also observed in the 1990s. These facts could be explained by the changing dynamics of water masses off the Peruvian coast. In fact, one of the authors of this paper observed a slightly different trend between SST at 5°S and 12°S. Also, different spatial patterns of water masses prevailing over periods of time have been observed (O. Moron, pers. comm.). From the mid-1970s to the mid-1980s, a strong prevalence of oceanic warm and high salinity waters inside the first 60 nmi offshore has been reported, influencing a significant part of the coast. This observation agrees with the period of low zooplankton abundance, anchoveta, and the high abundance of sardine, jack mackerel, and mackerel. Also, the recovery of anchoveta coincides with the years 1984 and 1985, when cold upwelled water prevailed along the coast. These dynamics probably explain the slight change in the seasonality of zooplankton volumes among decades. The changes of local dynamics of water masses are definitely related to changes over the whole Pacific basin, especially in the southern hemisphere (Clarke and Lebedev, 1999).
We thank several scientists from Instituto del Mar del Perú (IMARPE) for historical record information and Sonia Sánchez and Luis O. Duarte for discussions of an earlier version of this paper. Special thanks are extended to “Gordie” Swartzman (University of Washington) for his valuable comments and help in finishing this work during his visit to IMARPE. Monthly in situ SST data were contributed by the “Dirección de Hidrografia y Navegacion (DHN)” of the Peruvian Navy. Participation in the 3rd International Zooplankton Production Symposium, 20–23 May 2003, Gijón, Spain, was made possible with financial aid from Consejo Nacional de Ciencia y Tecnología del Perú (CONCYTEC) and the Eastern Pacific Consortium for Research on Global Change in Coastal and Oceanic Systems (EPCOR). Finally, we recognize the very helpful comments of the two referees.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}