





Abstract
North Sea herring (Clupea harengus) are managed as a single stock, but maintaining a diversity of spawning components is considered important. However, the total catch from each of these components cannot be estimated easily because the components mix during the summer feeding season. The spawning origin of herring is determined from patterns in the microstructure of the otolith core, from samples taken in the central and northern North Sea during summer of 2004–2007. The annual catch composition of Dutch vessels is determined within a statistical framework that takes account of the spatial patterns in mixing of spawning components and the classification success of the method. Mixing of components varied between years, with steep latitudinal gradients in compositions in some years, resulting in pronounced between-year differences in estimated catch compositions. Differences in lengths-at-age between spawning components, in particular of the 2000 year class, may have caused the observed between-year changes in mixing of components. Our results indicate that estimates of compositions change when assumptions of perfect spatial mixing and perfect classification are relaxed, and can be uncertain in particular as a result of misclassifications, and that it may not be appropriate to assume that ratios between components are constant through time.Bierman, S. M., Dickey-Collas, M., van Damme, C. J. G., van Overzee, H. M. J., Pennock-Vos, M. G., Tribuhl, S. V., and Clausen, L. A. W. 2010. Between-year variability in the mixing of North Sea herring spawning components leads to pronounced variation in the composition of the catch. – ICES Journal of Marine Science, 67: 885–896.
Introduction
North Sea herring (Clupea harengus) are assessed and managed as one unit or stock (ICES, 1965; Burd, 1985; Cushing, 1992). If only the genetic make-up of the stock is considered, this management strategy appears appropriate (Mariani et al., 2005; Reiss et al., 2009). However, because of the phenotypic plasticity of the species (Winters and Wheeler, 1996; McQuinn, 1997; Dickey-Collas et al., 2009), components of the stock (Heincke, 1898) have differences in growth, maturation, and recruitment patterns (Cushing, 1967; Almatar and Bailey, 1989; Hulme, 1995). The southern or Downs component (herring that spawn in December and January in the southern North Sea and eastern English Channel) leaves the spawning areas to feed in the central and northern North Sea in summer (Cushing and Bridger, 1966) with the other spawning components (Orkney/Shetland, Buchan, and Banks; see Figure 1). These components may experience different fishing pressure (ICES, 2006a) and may differ in their resilience to exploitation (Andersen et al., 1974; Burd, 1985). Maintaining local populations and a diversity of spawning components is thought to be an important target for the management of herring fisheries (Stephenson et al., 1999; McPherson et al., 2001; Secor et al., 2009).
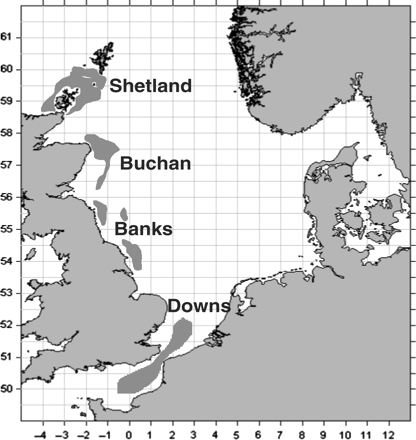
North Sea herring spawning components. Generalized major spawning grounds inferred from the presence of newly hatched larvae in the ICES herring larval survey, 1996–2003.
Attempts have been made to determine the spawning origin of North Sea herring from both commercial catches and surveys, using a variety of methods, including tagging, meristic characteristics, differences in length-at-age, and the supply of larvae (Cushing, 1958; Wood, 1959; ICES, 1965; Corten and van de Kamp, 1979; Burd and Hulme, 1984). However, the seasonal pattern in temperature development in the North Sea leads directly or indirectly (e.g. through the availability of food) to seasonal patterns in growth rates of larvae (Geffen, 2009), which in turn are shown in the microstructure or larval daily growth ring patterns in the otoliths of herring. Therefore, the components can be identified to their approximate spawning time by the otolith microstructure of the larval part of the otolith (Moksness and Fossum, 1991; Mosegaard and Madsen, 1996; Clausen et al., 2007), and this method has been used successfully in the Skagerrak and the Norwegian, Irish, and Celtic Seas (Stenevik et al., 1996; Brophy and Danilowicz, 2002).
We determined the spawning origin of herring, using the pattern in the microstructure of the otolith core, in samples taken from catches made in the central and northern North Sea during summers of 2004–2007. We estimated the annual catch composition of the herring taken by Dutch vessels, within a statistical framework that takes into account the spatial patterns in mixing of spawning components, along with the classification success of our method. Using mixed models, we tested for the presence of latitudinal gradients in proportions, and estimated, using variance components, whether extra-binomial variation was present in the observed proportions at the spatial level of the haul or of the grid cells. Within this statistical modelling framework, we were able to test whether there was evidence for (i) a spatial cline in the summer catches in the ratio between components, (ii) differences between years in this cline, and (iii) differences between years in the overall composition of the different spawner types in the landings. The suitability of the sampling programme and statistical modelling framework to estimate the spawning composition and the implications of our findings for management are discussed.
Methods
Samples
Samples of herring caught by Dutch vessels were collected from May to July of 2004–2007 (Figure 2). More than 20 samples per year (at least 25 fish per sample) were analysed from across the fishing area (Figure 2, Table 1). The spatial limit of samples was the area of the North Sea between 4°W and 5°E, and between 54 and 62°N (Figure 2). The total length of each herring was measured, and the sagittal otoliths removed and cleaned. One otolith was used for age determination (except for 2004), following standard ICES protocols, and the other to analyse the microstructure at the core. Left and right otoliths were used randomly for each analysis. Age–length keys of the whole population cannot be applied to the different spawner types. No specific autumn-spawner age–length key for 2004 was available, but a key for Downs herring for 2004 was applied to the winter-spawned fish, providing age information for winter spawners caught from 2004 to 2007 and for autumn spawners from 2005 to 2007. In 2006 and 2007, some samples from an acoustic survey were included to increase coverage (ICES, 2006b, 2007). Immature fish (<20 cm long) were discarded from the dataset.
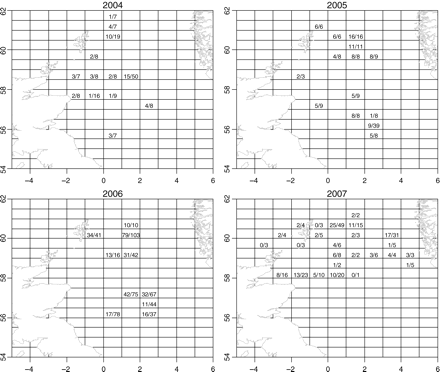
Samples of catches taken from the North Sea Dutch herring fleet from May to July, 2004–2007, and the observed proportions of autumn-spawned fish per ICES rectangle. Numerators are the numbers of autumn-spawned fish, and denominators are the total number of fish sampled. For some rectangles, more than one sample was available.
Summary, per year, of the number of samples (K), the total number of fish tested (N), the range in the number of fish per sample (range ni), the number of ICES rectangles within which at least one sample was taken (Q), the range in the number of samples per rectangle (range 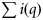 ), the range in the number of fish tested per rectangle (range
), the range in the number of fish tested per rectangle (range 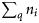 ), and the total observed numbers of autumn (
), and the total observed numbers of autumn (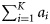 )-, winter (
)-, winter (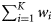 )-, and spring (
)-, and spring (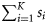 )-spawned fish.
)-spawned fish.
| Year | K | N | Range ni | Q | Range 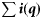 | Range 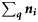 | 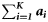 | 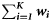 | 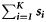 |
|---|---|---|---|---|---|---|---|---|---|
| 2004 | 20 | 162 | 7–10 | 13 | 1–6 | 7–50 | 51 | 103 | 8 |
| 2005 | 20 | 148 | 3–10 | 14 | 1–5 | 3–39 | 94 | 54 | 0 |
| 2006 | 27 | 513 | 5–42 | 10 | 1–6 | 10–103 | 285 | 228 | 0 |
| 2007 | 34 | 233 | 1–31 | 25 | 1–4 | 1–39 | 124 | 109 | 0 |
| Year | K | N | Range ni | Q | Range | Range | | | |
|---|---|---|---|---|---|---|---|---|---|
| 2004 | 20 | 162 | 7–10 | 13 | 1–6 | 7–50 | 51 | 103 | 8 |
| 2005 | 20 | 148 | 3–10 | 14 | 1–5 | 3–39 | 94 | 54 | 0 |
| 2006 | 27 | 513 | 5–42 | 10 | 1–6 | 10–103 | 285 | 228 | 0 |
| 2007 | 34 | 233 | 1–31 | 25 | 1–4 | 1–39 | 124 | 109 | 0 |
Summary, per year, of the number of samples (K), the total number of fish tested (N), the range in the number of fish per sample (range ni), the number of ICES rectangles within which at least one sample was taken (Q), the range in the number of samples per rectangle (range ), the range in the number of fish tested per rectangle (range ), and the total observed numbers of autumn ()-, winter ()-, and spring ()-spawned fish.
| Year | K | N | Range ni | Q | Range | Range | | | |
|---|---|---|---|---|---|---|---|---|---|
| 2004 | 20 | 162 | 7–10 | 13 | 1–6 | 7–50 | 51 | 103 | 8 |
| 2005 | 20 | 148 | 3–10 | 14 | 1–5 | 3–39 | 94 | 54 | 0 |
| 2006 | 27 | 513 | 5–42 | 10 | 1–6 | 10–103 | 285 | 228 | 0 |
| 2007 | 34 | 233 | 1–31 | 25 | 1–4 | 1–39 | 124 | 109 | 0 |
| Year | K | N | Range ni | Q | Range | Range | | | |
|---|---|---|---|---|---|---|---|---|---|
| 2004 | 20 | 162 | 7–10 | 13 | 1–6 | 7–50 | 51 | 103 | 8 |
| 2005 | 20 | 148 | 3–10 | 14 | 1–5 | 3–39 | 94 | 54 | 0 |
| 2006 | 27 | 513 | 5–42 | 10 | 1–6 | 10–103 | 285 | 228 | 0 |
| 2007 | 34 | 233 | 1–31 | 25 | 1–4 | 1–39 | 124 | 109 | 0 |
Estimates of the landings of herring were taken from the Dutch official logbook database (Visserij Registratie Informatie Systeem, VIRIS; Figure 3). Landings of herring are thought to be representative of catches because the proportion of discarded to landed herring from the Dutch fleet in the North Sea is small (<5%), and there is no size selectivity in the discards (Borges et al., 2008).
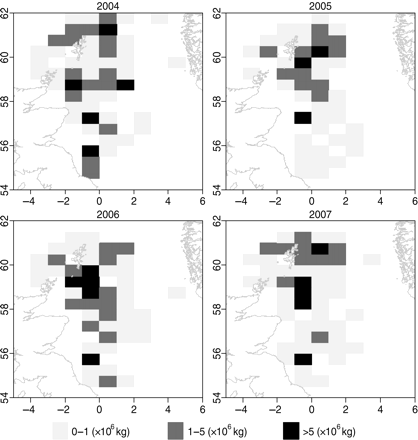
Herring landings into the Netherlands by ICES rectangle in the months May, June, and July of 2004–2007.
Determination of spawner type
To investigate the microstructure of the otolith core, the otoliths were mounted with the sulcus side up in thermoplastic resin (Buhler 40–8100) at 150°C, allowing for repeated relocation of the otolith for grinding and polishing on both sides. The otoliths were polished using grinding and polishing films with decreasing grain sizes from 30.0 to 0.3 µm to optimize the visual resolution to a focal plane through the otolith nucleus and a transect towards the edge large enough to see at least 100 day-rings. During the polishing process, the otolith was checked under a dissecting microscope to prevent it being overpolished. The ring pattern was read under a microscope using ×40–60 magnification. All otoliths were classified as autumn-, winter-, or spring-spawning types using the classification of Clausen et al. (2007).
Autumn spawners are characterized by primary increments, 2.5 µm wide, found 200 µm from the centre. All increments appeared to have close-to-constant widths. For winter spawners, the otolith increments gradually increase from ∼1 µm wide some 10 µm from the centre to 0.3 µm wide at a distance of 150 µm from the centre. The increase in increment widths accelerates at ∼200 µm from the centre. Spring spawners are highly variable spawner types characterized by wider and more optically dense increments than winter and autumn spawners. Early-hatched fish exhibit increments rapidly increasing from a width of 2 to 0.4 µm, and later-hatched fish have relatively wide increments of ∼4 µm already 20–40 µm from the nucleus.
Estimating the proportions of spawner types and the composition of the catch
For a given year, let ai, wi, and si be the number of autumn-, winter-, and spring-spawned fish, respectively, in sample i (i = 1,2, … , K, with K the total number of samples taken in that year). Further, let ni = ai = si = wi be the total number of fish in sample i, and 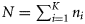 be the total number of fish tested in a year. We note that 2004 was the only year with spring-spawned fish in the samples (Table 1).
be the total number of fish tested in a year. We note that 2004 was the only year with spring-spawned fish in the samples (Table 1).
 or
or  . These estimates are obtained by multiplying the estimated proportions of autumn- and winter-spawned fish in rectangles,
. These estimates are obtained by multiplying the estimated proportions of autumn- and winter-spawned fish in rectangles, 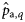 and
and 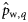 (with q = 1,2, … , Q, q being an indicator for rectangle and Q the total number of rectangles in a given year), with the total quantities of herring landed during May, June, and July in rectangles. For example, for
(with q = 1,2, … , Q, q being an indicator for rectangle and Q the total number of rectangles in a given year), with the total quantities of herring landed during May, June, and July in rectangles. For example, for 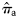 (similarly for
(similarly for 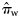 , see Table 1 and Figure 2),
, see Table 1 and Figure 2), 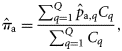
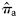 without using the catch data as
without using the catch data as 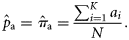
Approximate 95% confidence limits of this estimate are given by 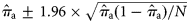 , using the normal approximation to the binomial distribution (because N is reasonably large in all years).
, using the normal approximation to the binomial distribution (because N is reasonably large in all years).
The data did not arise from a designed survey, resulting in spatial patterns in sampling effort such as differing numbers of fish tested per sample, spatio-temporal clustering of samples, and differences between years in the spatial extent of sampling (Figure 2, Table 1). Moreover, it is possible or even likely that autumn- and winter-spawned fish are not perfectly mixed throughout the whole area. Following from the spatial pattern of the sampling locations and exploratory analyses of the data (Figure 4), we distinguished three types of deviation from the perfect mixing situation.
There is evidence of broad-scale (across ICES rectangles) spatial gradients with latitude in the proportions of spawning types in the samples (Figure 4).
Perhaps fish of a particular spawning type will aggregate (e.g. in schools) at the spatial and temporal scale of the haul. This in turn may lead to a higher-than-expected (under the binomial distribution) variation in the proportions of autumn- and winter-spawned fish in samples (overdispersion). We refer to this as a sample effect.
Perhaps there is variation in the mixing of spawning types at intermediate spatial scales. Here, we take the ICES rectangle levels as our spatial unit because these rectangles also represent reasonably well the clustering of samples. We refer to this as a rectangle effect.
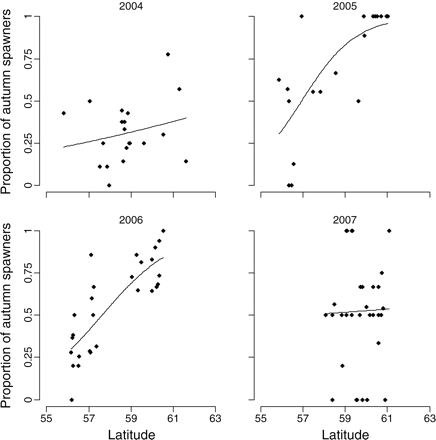
Sample proportions by sample latitude. The sample proportions were computed as the number of autumn-spawned fish over the total number of fish in each sample. The lines are fitted trends, with the latitude estimated using Equation (3) and the parameter estimates in Table 3.
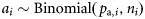
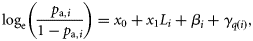
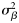 and
and 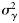 , respectively. Because the latitudes Li were scaled to have zero mean, parameter x0 provides an estimate of the proportion of autumn-spawned fish at the average latitude of the samples.
, respectively. Because the latitudes Li were scaled to have zero mean, parameter x0 provides an estimate of the proportion of autumn-spawned fish at the average latitude of the samples.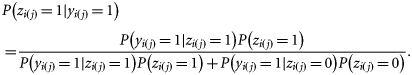
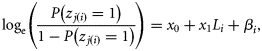
Estimates (and s.e.) of fixed effects (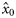 , intercept;
, intercept; 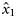 , slope for latitude) and variance components (
, slope for latitude) and variance components (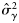 , variance component for rectangle;
, variance component for rectangle; 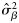 , variance component for haul) obtained by fitting the mixed model [Equations (3a) and (3b); models fitted using REML) to the proportions of autumn spawners.
, variance component for haul) obtained by fitting the mixed model [Equations (3a) and (3b); models fitted using REML) to the proportions of autumn spawners.
| Model parameter | 2004 | 2005 | 2006 | 2007 |
|---|---|---|---|---|
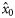 | −0.783 (0.254) | 1.243 (0.557) | 0.315 (0.170) | 0.130 (0.1383) |
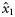 | 0.173 (0.215) | 2.013 (0.652) | 1.013 (0.180) | 0.028 (0.1186) |
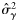 | 0.140 (0.350) | 0.188 (1.402) | 0.009 (0.120) | 0 |
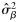 | 0.038 (0.280) | 2.161 (1.652) | 0.342 (0.208) | 0 |
| Model parameter | 2004 | 2005 | 2006 | 2007 |
|---|---|---|---|---|
| −0.783 (0.254) | 1.243 (0.557) | 0.315 (0.170) | 0.130 (0.1383) |
| 0.173 (0.215) | 2.013 (0.652) | 1.013 (0.180) | 0.028 (0.1186) |
| 0.140 (0.350) | 0.188 (1.402) | 0.009 (0.120) | 0 |
| 0.038 (0.280) | 2.161 (1.652) | 0.342 (0.208) | 0 |
Estimates (and s.e.) of fixed effects (, intercept; , slope for latitude) and variance components (, variance component for rectangle; , variance component for haul) obtained by fitting the mixed model [Equations (3a) and (3b); models fitted using REML) to the proportions of autumn spawners.
| Model parameter | 2004 | 2005 | 2006 | 2007 |
|---|---|---|---|---|
| −0.783 (0.254) | 1.243 (0.557) | 0.315 (0.170) | 0.130 (0.1383) |
| 0.173 (0.215) | 2.013 (0.652) | 1.013 (0.180) | 0.028 (0.1186) |
| 0.140 (0.350) | 0.188 (1.402) | 0.009 (0.120) | 0 |
| 0.038 (0.280) | 2.161 (1.652) | 0.342 (0.208) | 0 |
| Model parameter | 2004 | 2005 | 2006 | 2007 |
|---|---|---|---|---|
| −0.783 (0.254) | 1.243 (0.557) | 0.315 (0.170) | 0.130 (0.1383) |
| 0.173 (0.215) | 2.013 (0.652) | 1.013 (0.180) | 0.028 (0.1186) |
| 0.140 (0.350) | 0.188 (1.402) | 0.009 (0.120) | 0 |
| 0.038 (0.280) | 2.161 (1.652) | 0.342 (0.208) | 0 |
It is difficult to obtain good information on classification success, and because no single method can claim to be perfect, there is no golden standard. However, Clausen et al. (2007) compared the performance of the visual assignment method (as used here) on 0-group herring with a method in which hatching dates were estimated by enumerating numbers of day-rings (the back-tracked hatch season method). As the back-tracked method can be assumed to give precise estimates of hatching dates, we can use the results of this comparison (Table 4 of Clausen et al., 2007) to obtain estimates of the sensitivity and specificity of the visual assignment method. This yields estimates of sensitivity of P(yj(i) = 1|zi(j) = 1) = 90/100 = 0.90, and specificity of P(yj(i) = 0|zi(j) = 0) = 80/97 = 0.82. Here, we estimate the parameters in Equation (4b) below using two scenarios: (i) assuming perfect classification (sensitivity = specificity = 1) and (ii) allowing two scenarios for imperfect classification using sensitivity = specificity = 0.9 (an upper bound on classification success) and sensitivity = specificity = 0.82 (a lower bound on classification success).
We used the vaguely informative priors for the parameters of logistic models as proposed by Gelman et al. (2008): Student's t distributions with 7 degrees of freedom, and scale-parameters of 5 for the intercept (x0) and 2.5 for the slope for latitude [the covariate latitude was scaled to have zero mean and a standard deviation of 0.5 according to Gelman et al. (2008)], and a uniform prior between 0 and 15 for σβ.
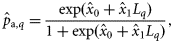
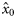 and
and 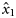 are simulated from their joint posterior distribution.
are simulated from their joint posterior distribution.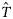 ) is then obtained by multiplying the estimated proportions by the landings per rectangle:
) is then obtained by multiplying the estimated proportions by the landings per rectangle: 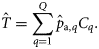
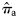 ) can be obtained from Equation (1).
) can be obtained from Equation (1).We note that 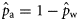 and
and 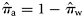 for the 2005, 2006, and 2007 data, because in those years there were no spring spawners in the sample. We implemented the Bayesian model [Equations (4a)–(4c)] under the assumption of perfect classification success also for the 2000 year class of each spawner type to estimate the proportions of the autumn (
for the 2005, 2006, and 2007 data, because in those years there were no spring spawners in the sample. We implemented the Bayesian model [Equations (4a)–(4c)] under the assumption of perfect classification success also for the 2000 year class of each spawner type to estimate the proportions of the autumn (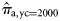 ) and winter spawners (
) and winter spawners (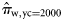 ) of this year class in the landings.
) of this year class in the landings.
Results
There was strong evidence of differences between years in the ratios of autumn to winter spawners in the landings (Tables 2 and 3; Figures 5–7). Small numbers of spring spawners were found in 2004 only, so we do not further consider the presence of spring spawners as part of the composition of the catch. In 2004 and 2007, there was little evidence of a spatial trend or of a sample or rectangle effect in the proportions of spawner types (Tables 2 and 3). Therefore, for those years, estimates of the composition of the catches obtained using simple averages [assumption of perfect mixture and perfect classification; Equation (2)] and using the model-based approach with perfect classification [Equations (4a)–(4c)] were similar (Figure 5). In contrast, there was strong evidence in 2005 and 2006 that the proportions of autumn spawners, both of all year classes combined and of the 2000 year class alone, increased with latitude (Figure 4; Tables 2 and 3). In addition, there was some evidence of a sample effect on top of this, as indicated by the relatively large estimates of the variance components 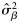 (Tables 2 and 3). Hence, in 2005 and 2006, the assumption of perfect mixing was clearly violated, and model-based estimates differed substantially from the estimates obtained under the assumption of perfect mixing (Figures 5 and 6). Relaxing the assumption of perfect classification resulted in shifts in the estimates of compositions towards the most dominant spawner type, and an increase in the uncertainty surrounding the estimates (Figures 5 and 6). With assumed values of sensitivity and specificity of 0.9, all models converged without a problem. However, with assumed values of sensitivity and specificity of 0.82, convergence was slow for the 2004 data, because the situation with all, or nearly all, fish being in truth winter spawners was not unlikely given the data (Figure 5). The spatial cline in proportions was steepest in 2005 (Table 3, Figure 4) and, because of the spatial distribution of the catches (Figure 3), resulted in the estimates of >80% (assuming perfect classification) and 86% (assuming sensitivity-specificity of 0.9) of the landed herring being autumn spawners after accounting for latitude and extrapolating proportions (Figures 5 and 6). This contrasted sharply with 2004, when ∼30% (assuming perfect classification) and 24% (assuming specificity = sensitivity = 0.9) of the landings consisted of autumn spawners. In 2007, there were almost equal catches of autumn and winter spawners. A large proportion of the total catches consisted of the year class that spawned in 2000, although, as expected, the numbers of that year class in the catch declined between 2004 and 2007 (Figures 6 and 7). The between-year variation in the estimated proportions of spawner types of the 2000 year class in the landings was similar to those of the landings of all year classes, with large proportions of winter spawners in 2004 and of autumn spawners in 2005, whereas almost equal proportions of each spawner type were landed in 2007 (cf. Figures 6 and 7). The signals for the other year classes over the short period of this study proved difficult to interpret.
(Tables 2 and 3). Hence, in 2005 and 2006, the assumption of perfect mixing was clearly violated, and model-based estimates differed substantially from the estimates obtained under the assumption of perfect mixing (Figures 5 and 6). Relaxing the assumption of perfect classification resulted in shifts in the estimates of compositions towards the most dominant spawner type, and an increase in the uncertainty surrounding the estimates (Figures 5 and 6). With assumed values of sensitivity and specificity of 0.9, all models converged without a problem. However, with assumed values of sensitivity and specificity of 0.82, convergence was slow for the 2004 data, because the situation with all, or nearly all, fish being in truth winter spawners was not unlikely given the data (Figure 5). The spatial cline in proportions was steepest in 2005 (Table 3, Figure 4) and, because of the spatial distribution of the catches (Figure 3), resulted in the estimates of >80% (assuming perfect classification) and 86% (assuming sensitivity-specificity of 0.9) of the landed herring being autumn spawners after accounting for latitude and extrapolating proportions (Figures 5 and 6). This contrasted sharply with 2004, when ∼30% (assuming perfect classification) and 24% (assuming specificity = sensitivity = 0.9) of the landings consisted of autumn spawners. In 2007, there were almost equal catches of autumn and winter spawners. A large proportion of the total catches consisted of the year class that spawned in 2000, although, as expected, the numbers of that year class in the catch declined between 2004 and 2007 (Figures 6 and 7). The between-year variation in the estimated proportions of spawner types of the 2000 year class in the landings was similar to those of the landings of all year classes, with large proportions of winter spawners in 2004 and of autumn spawners in 2005, whereas almost equal proportions of each spawner type were landed in 2007 (cf. Figures 6 and 7). The signals for the other year classes over the short period of this study proved difficult to interpret.
![Annual estimates (and 95% confidence limits) of the proportions of each spawning type in landings into the Netherlands, obtained under the assumptions of (i) perfect mixing and perfect classification [circles, Equation (2)], (ii) imperfect mixing (a latitudinal cline in proportions and a sample effect) and perfect classification [triangles, Equations (4a)–(4c) with sensitivity 1 and specificity 1], (iii) imperfect mixing and imperfect classification [squares; Equations (4a)–(4c) with both sensitivity and specificity of 0.9], and (iv) imperfect mixing and imperfect classification [diamonds; Equations (4a)–(4c) with sensitivity and specificity of 0.82]. Crosses indicate the proportions of spring spawners (no confidence limits given; only observed in 2004); filled symbols, proportions of autumn spawners; open symbols, proportions of winter spawners.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/icesjms/67/5/10.1093_icesjms_fsp300/1/m_fsp30005.gif?Expires=1716473573&Signature=HkkUCDIcoHj9PiG7QcXijF1Im4JrCs3kzKWKV8IemOZhLyJ3hW23cBwxYdM-cE1Nd4FjxySi9xAvbUiWNdFph6bNzr3EuatnqXjFPfbmRff2aHDr19q1mai4Gs97uBgwa4qyQn2kpgOiEkrcu7bugjx0BaXMcNamqK9T07J94ffS-gbRZZ8UgIn36lcDVN69BJZLVuyrJqvnLtixZ02zg-SqRL2ZSu5A3nB8I-gQZEcbFRiRMrSVRST9g4Iz7TdSkG6sZH0JzcdwD5~i4m23ZSFkZyQB2KnHOA-TSNKPnsK2Et5o1scFx8m73BXdDafYMezIKmVtHQT0tGCciX-Maw__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Annual estimates (and 95% confidence limits) of the proportions of each spawning type in landings into the Netherlands, obtained under the assumptions of (i) perfect mixing and perfect classification [circles, Equation (2)], (ii) imperfect mixing (a latitudinal cline in proportions and a sample effect) and perfect classification [triangles, Equations (4a)–(4c) with sensitivity 1 and specificity 1], (iii) imperfect mixing and imperfect classification [squares; Equations (4a)–(4c) with both sensitivity and specificity of 0.9], and (iv) imperfect mixing and imperfect classification [diamonds; Equations (4a)–(4c) with sensitivity and specificity of 0.82]. Crosses indicate the proportions of spring spawners (no confidence limits given; only observed in 2004); filled symbols, proportions of autumn spawners; open symbols, proportions of winter spawners.
![Estimated total quantities of autumn- and winter-spawned herring landed into the Netherlands [ in Equation (4c)], under three scenarios for classification success: (i) perfect classification with sensitivity and specificity of 1 (circles), (ii) imperfect classification with sensitivity and specificity of 0.9 (triangles), and (iii) imperfect classification with sensitivity and specificity of 0.82 (squares). Filled symbols, autumn spawners; open symbols, winter spawners.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/icesjms/67/5/10.1093_icesjms_fsp300/1/m_fsp30006.gif?Expires=1716473573&Signature=fkSKPcMguNJh-BYpM51pITqSaRkm65NazZfVGE5u6agljpBloZqEHdv7eg4BQubiFaTaoWSjMd2wrzJUnRC1YVEfonbRjkkZWcyeUFmDZDUW8YKcaem~WHkQ5n50B8a7Qevs7cR4e1I88gMGHLhiNkkzokFOi4t3yHE-YDhhyYxDqunVL3~W3Zhr3R84~BKWp0Q18axd6eHFYT7YPw9VyBb9lGXVIxobkroOOoZzNvJtdgt-Orok--F4oNotnTMPsVhp5C-XuwVB2ROjfBV694Ogxcq-mUi9Hc6eFx82H970C7xJyQZsMilF0HyvT7~HvRbPRmzrL~hEcuVvPfRQeQ__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Estimated total quantities of autumn- and winter-spawned herring landed into the Netherlands [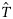 in Equation (4c)], under three scenarios for classification success: (i) perfect classification with sensitivity and specificity of 1 (circles), (ii) imperfect classification with sensitivity and specificity of 0.9 (triangles), and (iii) imperfect classification with sensitivity and specificity of 0.82 (squares). Filled symbols, autumn spawners; open symbols, winter spawners.
in Equation (4c)], under three scenarios for classification success: (i) perfect classification with sensitivity and specificity of 1 (circles), (ii) imperfect classification with sensitivity and specificity of 0.9 (triangles), and (iii) imperfect classification with sensitivity and specificity of 0.82 (squares). Filled symbols, autumn spawners; open symbols, winter spawners.
![Estimated total quantities of autumn- and winter-spawned herring of the 2000 year class landed into the Netherlands [ in Equation (4c)]. Estimates were obtained by multiplying estimated proportions [Equations (4a)–(4c)] in quadrants with the landings reported from those quadrants. Open circles, autumn spawners; open triangles, winter spawners.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/icesjms/67/5/10.1093_icesjms_fsp300/1/m_fsp30007.gif?Expires=1716473573&Signature=neE9F2wyaK4hQgCKT37L~I~hZChqNM5x1BJlFytPwyJm-oIRbUb-YoSdbiPid2jUZr-a5xXFRKxjAnAT2~1Og6LZZGxb3L19fT~HZiviw5YzlIzETy74mUuVS5SovlZ9SLbkF6a-YEQFtKBhho5nJYYBgkOjlPiKbXOaf8yQ-fTJIlK0IfFD73P42IEA1PGwEyvRHN1eXxukRk3J23TSXvr1AXwRKycqhEgDLjutTilWPzlZvzhJDJEgQMSInQGNE0oNUq1zhJEpdJp~JknIB4B05VelnM57yUpCwpvQznqgYepVUmpnvHS3FgUMOaryliEc7LcLyERo2ca90JPrqg__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Estimated total quantities of autumn- and winter-spawned herring of the 2000 year class landed into the Netherlands [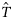 in Equation (4c)]. Estimates were obtained by multiplying estimated proportions [Equations (4a)–(4c)] in quadrants with the landings reported from those quadrants. Open circles, autumn spawners; open triangles, winter spawners.
in Equation (4c)]. Estimates were obtained by multiplying estimated proportions [Equations (4a)–(4c)] in quadrants with the landings reported from those quadrants. Open circles, autumn spawners; open triangles, winter spawners.
Means (and s.d.) of the posterior distributions of model parameters (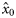 , intercepts;
, intercepts; 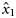 , slopes;
, slopes; 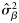 , variance component for samples) obtained by fitting the Bayesian model [see Equations (4a) and (4b)] to observed proportions of autumn spawners (to estimate
, variance component for samples) obtained by fitting the Bayesian model [see Equations (4a) and (4b)] to observed proportions of autumn spawners (to estimate 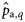 ), winter spawners (to estimate
), winter spawners (to estimate 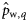 ), autumn spawners of the 2000 year class (to estimate
), autumn spawners of the 2000 year class (to estimate 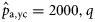 ), and winter spawners of the 2000 year class (to estimate
), and winter spawners of the 2000 year class (to estimate 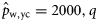 ).
).
| Estimated proportion | Model parameter | Value per year | |||
|---|---|---|---|---|---|
| 2004 | 2005 | 2006 | 2007 | ||
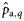 | 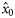 | −0.84 (0.22) | 1.59 (0.77) | 0.33 (0.17) | 0.13 (0.15) |
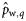 | 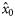 | 0.60 (0.20) | −1.59a (0.77) | −0.33a (0.17) | −0.13a (0.15) |
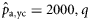 | 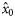 | n/a | −0.52 (0.24) | −1.47 (0.20) | −2.02 (0.34) |
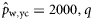 | 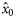 | n/a | −2.58 (0.54) | −2.35 (0.31) | −2.28 (0.27) |
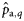 | 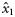 | 0.41 (0.40) | 4.56 (1.41) | 2.01 (0.35) | 0.08 (0.32) |
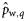 | 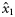 | −0.53 (0.39) | −4.56a(1.41) | −2.01a(0.35) | −0.08a (0.32) |
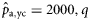 | 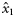 | n/a | 1.211 (0.47) | 0.76 (0.25) | 0.47 (0.63) |
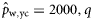 | 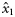 | n/a | −3.29 (1.01) | −1.31 (0.56) | 0.64 (0.51) |
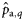 | 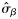 | 0.46 (0.29) | 2.42 (0.92) | 0.69 (0.20) | 0.26 (0.22) |
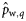 | 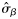 | 0.36 (0.26) | 2.42a (0.92) | 0.69a (0.20) | 0.26a (0.22) |
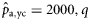 | 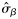 | n/a | 0.57 (0.36) | 0.73 (0.25) | 0.81 (0.63) |
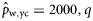 | 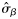 | n/a | 0.80 (0.48) | 1.07 (0.34) | 0.33 (0.29) |
| Estimated proportion | Model parameter | Value per year | |||
|---|---|---|---|---|---|
| 2004 | 2005 | 2006 | 2007 | ||
| | −0.84 (0.22) | 1.59 (0.77) | 0.33 (0.17) | 0.13 (0.15) |
| | 0.60 (0.20) | −1.59a (0.77) | −0.33a (0.17) | −0.13a (0.15) |
| | n/a | −0.52 (0.24) | −1.47 (0.20) | −2.02 (0.34) |
| | n/a | −2.58 (0.54) | −2.35 (0.31) | −2.28 (0.27) |
| | 0.41 (0.40) | 4.56 (1.41) | 2.01 (0.35) | 0.08 (0.32) |
| | −0.53 (0.39) | −4.56a(1.41) | −2.01a(0.35) | −0.08a (0.32) |
| | n/a | 1.211 (0.47) | 0.76 (0.25) | 0.47 (0.63) |
| | n/a | −3.29 (1.01) | −1.31 (0.56) | 0.64 (0.51) |
| | 0.46 (0.29) | 2.42 (0.92) | 0.69 (0.20) | 0.26 (0.22) |
| | 0.36 (0.26) | 2.42a (0.92) | 0.69a (0.20) | 0.26a (0.22) |
| | n/a | 0.57 (0.36) | 0.73 (0.25) | 0.81 (0.63) |
| | n/a | 0.80 (0.48) | 1.07 (0.34) | 0.33 (0.29) |
As the herring in the 2004 samples were not subjected to age determination, no estimates were obtained for 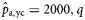 and
and 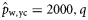 for that year (indicated by n/a). Slopes that were estimated to be significantly different from zero (if the 2.5 and 97.5% quantiles of the posterior distributions did not include zero) are shown emboldened.
for that year (indicated by n/a). Slopes that were estimated to be significantly different from zero (if the 2.5 and 97.5% quantiles of the posterior distributions did not include zero) are shown emboldened.
aIn the years 2005, 2006, and 2007, no spring spawners were observed, so 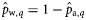 . Therefore, values for
. Therefore, values for 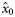 and
and 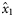 are of equal magnitude but of opposite sign, whereas values for
are of equal magnitude but of opposite sign, whereas values for 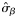 are identical.
are identical.
Means (and s.d.) of the posterior distributions of model parameters (, intercepts; , slopes; , variance component for samples) obtained by fitting the Bayesian model [see Equations (4a) and (4b)] to observed proportions of autumn spawners (to estimate ), winter spawners (to estimate ), autumn spawners of the 2000 year class (to estimate ), and winter spawners of the 2000 year class (to estimate ).
| Estimated proportion | Model parameter | Value per year | |||
|---|---|---|---|---|---|
| 2004 | 2005 | 2006 | 2007 | ||
| | −0.84 (0.22) | 1.59 (0.77) | 0.33 (0.17) | 0.13 (0.15) |
| | 0.60 (0.20) | −1.59a (0.77) | −0.33a (0.17) | −0.13a (0.15) |
| | n/a | −0.52 (0.24) | −1.47 (0.20) | −2.02 (0.34) |
| | n/a | −2.58 (0.54) | −2.35 (0.31) | −2.28 (0.27) |
| | 0.41 (0.40) | 4.56 (1.41) | 2.01 (0.35) | 0.08 (0.32) |
| | −0.53 (0.39) | −4.56a(1.41) | −2.01a(0.35) | −0.08a (0.32) |
| | n/a | 1.211 (0.47) | 0.76 (0.25) | 0.47 (0.63) |
| | n/a | −3.29 (1.01) | −1.31 (0.56) | 0.64 (0.51) |
| | 0.46 (0.29) | 2.42 (0.92) | 0.69 (0.20) | 0.26 (0.22) |
| | 0.36 (0.26) | 2.42a (0.92) | 0.69a (0.20) | 0.26a (0.22) |
| | n/a | 0.57 (0.36) | 0.73 (0.25) | 0.81 (0.63) |
| | n/a | 0.80 (0.48) | 1.07 (0.34) | 0.33 (0.29) |
| Estimated proportion | Model parameter | Value per year | |||
|---|---|---|---|---|---|
| 2004 | 2005 | 2006 | 2007 | ||
| | −0.84 (0.22) | 1.59 (0.77) | 0.33 (0.17) | 0.13 (0.15) |
| | 0.60 (0.20) | −1.59a (0.77) | −0.33a (0.17) | −0.13a (0.15) |
| | n/a | −0.52 (0.24) | −1.47 (0.20) | −2.02 (0.34) |
| | n/a | −2.58 (0.54) | −2.35 (0.31) | −2.28 (0.27) |
| | 0.41 (0.40) | 4.56 (1.41) | 2.01 (0.35) | 0.08 (0.32) |
| | −0.53 (0.39) | −4.56a(1.41) | −2.01a(0.35) | −0.08a (0.32) |
| | n/a | 1.211 (0.47) | 0.76 (0.25) | 0.47 (0.63) |
| | n/a | −3.29 (1.01) | −1.31 (0.56) | 0.64 (0.51) |
| | 0.46 (0.29) | 2.42 (0.92) | 0.69 (0.20) | 0.26 (0.22) |
| | 0.36 (0.26) | 2.42a (0.92) | 0.69a (0.20) | 0.26a (0.22) |
| | n/a | 0.57 (0.36) | 0.73 (0.25) | 0.81 (0.63) |
| | n/a | 0.80 (0.48) | 1.07 (0.34) | 0.33 (0.29) |
As the herring in the 2004 samples were not subjected to age determination, no estimates were obtained for and for that year (indicated by n/a). Slopes that were estimated to be significantly different from zero (if the 2.5 and 97.5% quantiles of the posterior distributions did not include zero) are shown emboldened.
aIn the years 2005, 2006, and 2007, no spring spawners were observed, so . Therefore, values for and are of equal magnitude but of opposite sign, whereas values for are identical.
In 2005 and 2006, the mean lengths-at-age of winter-spawned fish were significantly smaller (ANOVA; difference of 1.18 cm, p < 0.001, F-test with 1 and 139 d.f. in 2005, and a difference of 1.03 cm, p < 0.001, F-test with 1 and 114 d.f. in 2006) than those of autumn spawners (Figure 8). The 2000 year class grew more slowly than the other year classes (ICES, 2006a) and could be tracked from 2005 through 2007 (Figure 8). The differences in mean length-at-age were greatest for fish of the 2000 year class in 2005, with a difference of 1.44 cm, resulting in 4-year-old fish in our samples from that year having a slightly (although not significantly) higher mean length than the 5-year olds in the same sample (Figure 8). The lengths of herring of both spawner types in our samples increased with latitude in all years, with an average length of ∼28 cm at a latitude of 61°, compared with ∼23–25 cm at 57° latitude (Figure 9).
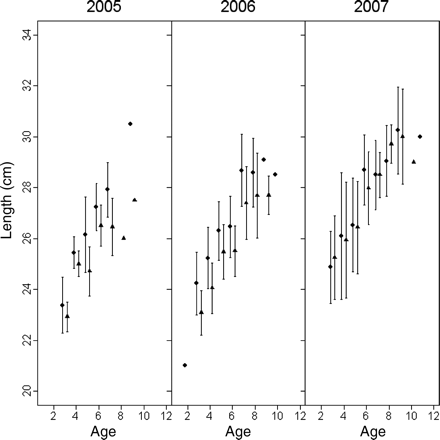
Mean (±s.e.) lengths-at-age of autumn- and winter-spawned fish in the samples obtained from 2005 to 2007 (fish in the 2004 samples were not subjected to age determination).
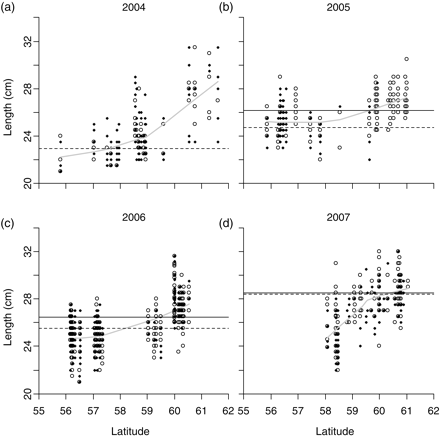
Lengths of winter- (dots) and autumn- (open circles) spawned fish observed in the samples, plotted against the latitude of the haul from which the samples were taken. Superimposed on the graphs are the mean lengths of autumn- (horizontal lines) and winter- (dashed horizontal lines) spawned fish of the 2000 year class. No estimates were available for 4-year-old autumn-spawned herring in 2004, because fish from that year were not subjected to age determination. The mean length of 4-year-old winter-spawned herring in 2004 was estimated using an age–length key of winter-spawned fish obtained from market sampling (see the Material and Methods section). To aid visual interpretation, smooth lines (obtained using a scatterplot smoother) through all the points are given as estimates of mean lengths at latitude.
Discussion
We estimated pronounced differences between years in the ratios of the proportions of winter and autumn spawners in the landings of herring into the Netherlands from the summer fishery on herring in the North Sea. We note that from 2004 to 2007, there were very few spring-spawned herring in our samples, which shows that proportions of Norwegian, western Baltic, and coastal spring-spawning herring were low among the summer feeding aggregations of herring in that part of the North Sea sampled here. Although this study concentrated on the Dutch fleet, the German, English, and part of the French fleets operate in the same way, so it may be assumed that the catch compositions for these fleets would have been similar. Other fleets (Danish, Norwegian, Scottish, etc.) could be expected to have had different catch compositions in 2005 and 2006 (the years in which a latitudinal cline in proportions of spawning types in the North Sea was found) if their spatial (in particular latitudinal) distribution of catches differed.
The results of this study showed that with the current methodology, the composition of the catch can be estimated with relatively high precision in some years, but with considerable uncertainty in other years. In any case, significant differences can be detected between years in the proportions of spawner types in the landings of herring (Figure 6). Our results clearly demonstrate that both spatial patterns in the mixing of components, and potential imperfect classification, need to be taken into account when estimating the composition. The main source of uncertainty in our estimates is that of the success of the classification method, in particular the values for sensitivity and specificity. The net effect of relaxing the assumption of perfect classification is that estimates of proportions of the dominant spawner type increase. This is logical, because most of the misclassified fish will be of the most dominant spawner type. For that reason, estimates change only little when proportions are close to 0.5 (such as in 2007; Figure 5), but most when one of the components is dominant (such as in 2004; Figure 5). Although autumn spawners dominated in 2005, the 2005 estimates were less affected because the proportions of winter spawners were still estimated to be high in the southern North Sea. Therefore, we believe that the precision of the estimates of herring spawning components will increase most by obtaining better information on classification success. Longer time-series and further analyses are needed to investigate whether the precision of the estimates of proportions of spawner types are high enough to provide useful information with which to follow cohorts. Moreover, our classification methodology relies on differences in increment width between spawner types. Perhaps these are partly influenced by interannual variation in growth conditions, such as food availability and temperature. More research is therefore needed to assess interannual variability in these interval widths.
An important finding of this study is that the mixing of the subcomponents can vary greatly between years, with steep latitudinal gradients in the composition of samples in some years, but not in others. Although the increasing proportions of autumn-spawned fish with latitude, as found in 2005 and 2006, agree with the notion that winter-spawned fish tend to remain more to the south of the summer feeding area than autumn-spawned fish (Cushing and Bridger, 1966; Burd, 1978), the results from 2004 and 2007 suggest that the spatial distribution of both components overlapped in those years. Over the period studied, but in particular in 2004 and 2005, the year class that spawned in 2000 dominated the catches, and it had been hypothesized that this year class was dominated by winter-spawned fish (ICES, 2006a). Our results support this hypothesis (Figures 5 and 6). There was no evidence that the increase in numbers of autumn spawners between 2004 and 2005 was supported by the appearance of a strong new autumn year class nor by the disappearance of a particular winter year class. We therefore hypothesize that, although the relatively large catch of winter spawners in 2004 resulted in a decrease in the overall proportion of winter spawners (in particular of the 2000 year class) in the North Sea, the coincidence of a latitudinal gradient in catches and mixing of the spawning types resulted in the large proportions of autumn spawners in the 2005 catch.
Winter-spawned herring in our samples, in particular those of the 2000 year class, were smaller at age than autumn-spawned herring. ICES assumes that full selectivity of North Sea herring is at age 5 (described by ICES, 2006a, as 4 winter rings). Therefore, perhaps the relative catchability of winter-spawned herring aged 4 was lower in 2004. Because of the large proportions of winter-spawned herring of the 2000 year class in 2004, we do not believe that larger autumn-spawned fish entered the fishery earlier by migrating earlier to the summer feeding areas. Instead, our findings suggest that, in 2004, neither winter- nor autumn-spawned fish of the 2000 year class were large enough to migrate farther north in large numbers during the feeding season (Figures 8 and 9). Our hypothesis is that this resulted in relatively good spatial overlap, and hence good mixing, of both components in the central and southern North Sea. In contrast, in 2005, herring of the autumn-spawned 2000 year class had grown large enough to migrate farther north in relatively larger numbers, whereas this was not the case for the winter spawners, resulting in the observed steep latitudinal gradient in the proportions. The same scenario applied, but to a lesser extent, in 2006, when the difference in mean length-at-age between winter and autumn spawners had decreased. By 2007, there was no evidence that winter spawners of the 2000 year class were smaller than their autumn-spawning counterparts, and herring of both spawner types were on average large enough to migrate farther north in relatively large numbers, resulting again in good mixing of both components. Other studies have suggested that migration distance in herring can be size-dependent (Slotte, 1999; Slotte and Fiksen, 2000). Therefore, we believe that the observed differences in growth rates between autumn- and winter-spawned fish, in particular of the strong but smaller-in-size 2000 year class, in combination with size-dependent migration patterns, underlie the observed between-year differences in the spatial overlap of the subcomponents.
We note that this study does not show that the fish caught in summer were actual autumn or winter spawners, but that they came from spawning events in autumn or winter. It is often assumed that most herring exhibit spawning fidelity in space and time. It is likely that some fish are vagrants or migrants from other spawning times or places (see McQuinn, 1997; Brophy and Danilowicz, 2003), but this is always assumed to be a small proportion of the population, unless there is a really strong year class. A linked study over the period 2003–2006 showed that all the herring caught by the Dutch fleet on the Downs herring spawning aggregations were winter-spawned (22 samples with 100% winter-spawned fish; ICES, 2005), so it is likely that our estimates are similar to the proportions of fish that are active autumn or winter spawners.
Downs herring (herring that spawn in December and January in the southern North Sea and eastern English Channel) feed in summer north of 53°30′N, and the regional TAC was designed to protect the migration to spawn and the spawning aggregations themselves. Currently, North Sea herring are managed with area-based total allowable catches (TACs), which apply a fixed ratio of the total North Sea TAC between areas (89% north of 53°30′N, 11% south of that line) to protect specific components of the stock. This management strategy was developed to protect the southern or Downs component when spawning (Figure 1). Downs herring leave the spawning areas to feed in the central North Sea in summer (Cushing and Bridger, 1966) with the other spawning components (Orkney/Shetland, Buchan, and Banks). Given a regional TAC, the quantity of each spawning type caught will be directly proportional to its ratio with the other component in the region. This could act as a buffer in that, if one of the subcomponents is small, relatively few fish of that subcomponent will be caught if they mix fully with the other subcomponent. However, if either subcomponent is small and does not mix well with the other subcomponent, that outcome is not certain. For example, if winter-spawned herring tend not to migrate as far north in large numbers as autumn-spawned herring in a particular year, almost all the fish caught in the north in that year will be autumn-spawned, regardless of the relative strengths of the two subcomponents. Each component shows different recruitment patterns (also see Winters and Wheeler, 1987; Myers, 2001), and this study clearly shows that spatial patterns in mixing of the components vary greatly between years. Given that the fishery exploits mixed aggregations of the spawning components, single-species stock assessments (such as currently in use for North Sea herring; Simmonds, 2009) will not accurately detect high rates of exploitation of individual components (Kell et al., 2009). Therefore, a management approach that is precautionary for the stock as a whole may not be precautionary for either spawning component individually. To design a more effective management strategy, it is therefore crucial that the mixing of stocks is well understood (Kell et al., 2009). Here, we have started to explore the possibilities and difficulties in utilizing the available information on herring spawning types in the central and northern North Sea to start monitoring the dynamics of mixing.
Acknowledgements
This research forms part of the strategic research programme “Sustainable spatial development of ecosystems, landscapes, seas and regions”, funded by the Dutch Ministry of Agriculture, Nature Conservation and Food Quality. We thank Jan Beintema and Yolande Jongejans for their help in collecting and reading otoliths, and the crews of the ships from which samples were taken for their assistance. Finally, we thank Beatriz Roel and an anonymous reviewer for their comments which greatly improved the manuscript.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}