





Abstract
European skate landings have traditionally been reported under a generic landing category, because of problems with species identification. To address this data deficiency, the ICES Working Group on Elasmobranch Fishes compiled conversion factors, including the relationships between different body measurements, for the main elasmobranch species. Size conversion factors for six common NE Atlantic skate species, Leucoraja naevus, Raja brachyura, R. clavata, R. miraletus, R. montagui, and R. undulata are compiled, and the capability of morphometric data to assist species discrimination is evaluated, highlighting the case of similar species such as R. brachyura and R. montagui. The estimated size conversion factors displayed some variability between areas and sexes for most species, the allometric relationship between weight and total length did not differ significantly between sexes, and some morphometric ratios proved adequate in discriminating between rajid species (misclassification error 0.12). Leucoraja naevus was fully discriminated from the remaining species. Species with a similar dorsal colour, e.g. R. brachyura and R. montagui, showed good discrimination based on their morphometry, with just 6–11% misclassification between the two.Serra-Pereira, B., Farias, I., Moura, T., Gordo, L. S., Santos, M., and Figueiredo, I. 2010. Morphometric ratios of six commercially landed species of skate from the Portuguese continental shelf, and their utility for identification. – ICES Journal of Marine Science, 67: 1596–1603.
Introduction
Skates (order Rajiformes) are one of the most speciose elasmobranch orders and include at least 27 genera and more than 245 species (Ebert and Compagno, 2007). They are found in all oceans, from shallow coastal waters to abyssal regions. Of those inhabiting the shelf and upper slope, some live on soft bottom, others on coarser substrata (Stehmann and Bürkel, 1984; Ebert and Compagno, 2007). Skates are important elements of the marine biodiversity, but they are highly vulnerable to commercial exploitation. In 2006, skate catches amounted for ∼59% of the total reported landings (by weight) of elasmobranchs in the NE Atlantic (FAO, 2007).
The external morphology and colour of skates are, in many cases, sufficient to discriminate between species inhabiting the NE Atlantic. Nevertheless, for commercial reasons, the different species are often landed under a generic landing category (Dulvy et al., 2000; ICES, 2007). During a pilot sampling programme conducted at ports in mainland Portugal, eight species of skate were identified: cuckoo ray Leucoraja naevus, blonde ray Raja brachyura, thornback ray Raja clavata, small-eyed ray Raja microocellata, brown ray Raja miraletus, spotted ray Raja montagui, undulate ray Raja undulata, and bottlenosed skate Rostroraja alba (Bordalo-Machado et al., 2004). In addition, three other species, which are either rare or absent from landings, have been described for this geographic area: longnosed skate Dipturus oxyrinchus, sandy ray Leucoraja circularis (Figueiredo et al., 2007), and Iberian pigmy skate Neoraja iberica (Stehmann et al., 2008). The present work focuses on six of the most common and widely studied species of this assemblage (L. naevus, R. brachyura, R. clavata, R. montagui, R. miraletus, and R. undulata). Identification keys are available for all six (e.g. Stehmann and Bürkel, 1984) to allow for their correct identification. Although the typical specimens of L. naevus, R. miraletus, and R. undulata are difficult to misidentify, because of their unique dorsal colouration, there may be problems distinguishing between the other three, mainly because of the remarkable variability within R. clavata, and the similarity between R. montagui and R. brachyura. Leucoraja naevus, R. miraletus, and R. montagui are small species that reach a maximum length of ∼70 cm (Holden, 1972; Du Buit, 1975), maturing at lengths of ∼60 cm (Walker, 1999). All three are potentially more resilient to fishing pressure than larger species, such as R. brachyura, R. clavata, and R. undulata, which can reach lengths of >100 cm (Holden, 1972; Moura et al., 2007; Serra-Pereira et al., 2008).
One of the terms of reference for the 2007 ICES Working Group on Elasmobranch Fishes (WGEF) referred to the need to compile conversion factors for elasmobranch species, but little information was made available (ICES, 2007). Conversion factors are commonly used to estimate values from one or more known body measurements. Therefore, morphometric conversions are particularly helpful when, for example, a specimen is damaged, or when dealing with commercially preprocessed specimens (e.g. wings only, tail off, head off, or a combination of these), in which not all morphometric traits can be measured. As conversion factors differ between species, they may also serve as a tool for species identification, especially for persons with limited taxonomic expertise, or for accurate identification of problematic specimens. This is particularly important at landing ports where fish need to be identified on site.
The present study aimed to (i) estimate relationships between different body measurements (i.e. size conversion factors) to increase the information available for six common NE Atlantic skate species in Portuguese waters, and (ii) investigate the ability of these measurements to discriminate between species to provide additional tools for assisting in the identification of skates on the Portuguese shelf and in other parts of the NE Atlantic.
Material and methods
Specimens were sampled monthly between February 2001 and June 2008, from commercial landings in three ports along the Portuguese continental coast: Matosinhos in the north, Peniche in the centre, and Portimão in the south (Figure 1). From these ports, skates are caught in a variety of gears, including trammel- and gillnets, longlines, trawls, and traps.
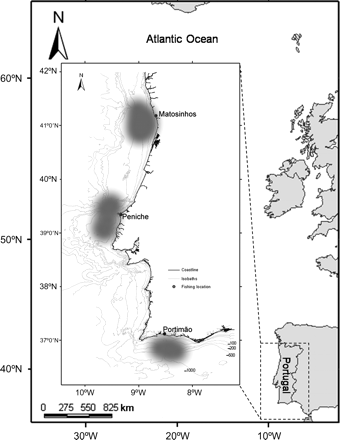
Map of the NE Atlantic with details of the study location off Portugal. The landing ports (Matosinhos, Peniche, and Portimão) and the main fishing areas (shaded grey) are identified, and bathymetric contours (isobaths) are provided in metres.
For each boat sampled, a subsample of the total skate landings was selected randomly. For each specimen, the species was identified and the following size measurements were recorded to the nearest millimetre (mm; see Figure 2): total length (TL), disc width (DW), disc length (DL), and tail length (CL). Although all the measurements are shown in Figure 2 on the ventral view of a skate, TL, DW, and DL were measured on linear axes under the individual, as viewed from the dorsal side. DL and CL were only available for specimens landed in Peniche. Specimens were also sexed and weighed (total weight, TW, and gutted weight, gW) to an accuracy of 1–10 g.
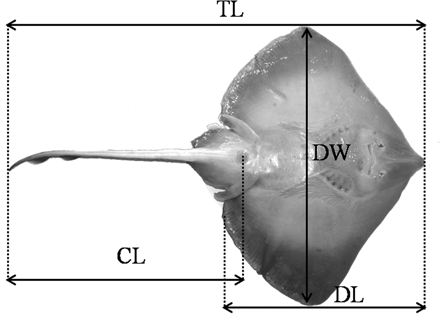
Measurements recorded to the nearest 1 mm on linear axes for each fish: total length (distance from the tip of the snout to the end of the tail, TL), disc width (maximum distance between the wing tips, DW), disc length (distance from the tip of the snout to the posterior edge of the disc, DL), and tail length (distance from the cloaca to the end of the tail, CL).
Data analysis
For each species, an exploratory analysis of morphometric ratios (DW:TL, DL:TL, CL:TL, and DL:DW) was performed by sex and area (landing port), whenever there were sufficient data available for two variables. Raja miraletus from the north and centre and L. naevus and R. undulata from the south were not considered for analysis owing to the small sample sizes by sex (n < 10). No R. brachyura were available in samples from the south.
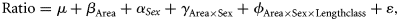
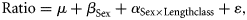
A stepwise algorithm was used to select the factors to be included in the model. Model selection was based on the Akaike Information Criterion (AIC), according to which, competing models may be ranked for a given dataset, and the one with the lowest AIC value is considered to be the most appropriate (Venables and Ripley, 2002).
Based on the statistical significance of the adjustments above, two procedures were used to estimate the conversion factors: (i) if the model selected explained >50% of the total variance, the size conversion factor was estimated from the combination of the different levels of the factors with a significant effect, using Equations (1) and (2); (ii) if the model captured only a small part of the total variance (<50%), the conversion factors were estimated by adjusting a linear model to the pairs of measurements under analysis, without considering the effect of the previous factors (sex, area, and TL class). The least-squares method was used to estimate the parameters of the expressions TL ∼ a DW + b, TL ∼ a CL + b, TL ∼ a DW + b, and DW ∼ a DL + b, where a is the intercept and b the slope.
The allometric relationships between TW and TL and between gW and TL were adjusted, and the parameters estimated by a non-linear least-squares method (Venables and Ripley, 2002): TW ∼ a TLb and gW ∼ a TLb, where a is the initial growth coefficient or condition factor and b represents the growth rate. Likelihood-ratio tests (Draper and Smith, 1981) were applied to compare the parameter estimates between sexes for the two length–weight relationships.
A flexible discriminant analysis (FDA; Hastie et al., 1994) was applied to investigate whether a simple combination of morphometric ratios was adequate to discriminate between species. All available species were used in these analyses. The FDA is a method for multigroup classification, and rules are built to predict the class membership of an item (species) based on several predictors, in this case the morphometric ratios. The morphometric ratios used as body-shape descriptors were: DW:TL (whole body shape), DL:DW (disc shape), and CL:DL (relative tail size).
Results
In all, 2009 fish were sampled, and Table 1 summarizes the number sampled by species, sex and geographical area, and their size ranges.
Summary of the data available by species, sex, and area, provided as ranges for each morphometric measurement.
| Species | Sex | Area | n | TL | TW | gW (g) | DW (mm) | DL (mm) | CL (mm) |
|---|---|---|---|---|---|---|---|---|---|
| JAI | F | South | 16 | 329–488 | – | – | 228–336 | – | – |
| M | South | 22 | 313–492 | – | – | 215–310 | – | – | |
| RJC | F | North | 150 | 420–905 | 200–7 750 | – | 305–608 | – | – |
| Centre | 155 | 489–934 | 623–6 710 | 585–5 810 | 345–669 | 248–489 | 269–464 | ||
| South | 22 | 362–793 | 900–3 800 | – | 263–588 | – | – | ||
| M | North | 133 | 410–875 | 450–4 140 | – | 295–562 | – | – | |
| Centre | 80 | 458–870 | 540–3 870 | 480–3 435 | 321–565 | 226–428 | 260–443 | ||
| South | 30 | 315–850 | 1 000–3 200 | – | 212–547 | – | – | ||
| RJH | F | North | 68 | 380–975 | 400–8 520 | – | 297–715 | – | – |
| Centre | 148 | 376–1 061 | 304–10 680 | 283–9 160 | 268–771 | 205–588 | 230–514 | ||
| M | North | 67 | 389–1 005 | 400–8 700 | – | 284–723 | – | – | |
| Centre | 114 | 412–1 005 | 421–7 290 | 387–6 404 | 292–692 | 240–554 | 260–495 | ||
| RJM | F | North | 33 | 368–702 | 270–2 340 | – | 257–459 | – | – |
| Centre | 185 | 390–702 | 388–2 583 | 360–2 306 | 285–483 | 212–373 | 240–346 | ||
| South | 18 | 331–575 | – | – | 235–412 | – | – | ||
| M | North | 23 | 412–651 | 450–1 980 | – | 281–430 | – | – | |
| Centre | 97 | 408–612 | 455–1 575 | 420–1 450 | 292–412 | 209–319 | 226–336 | ||
| South | 20 | 366–522 | – | – | 253–354 | – | – | ||
| RJN | F | North | 23 | 497–718 | 700–2 460 | – | 270–409 | – | – |
| Centre | 224 | 448–691 | 460–2 135 | 435–1 975 | 249–399 | 215–340 | 250–353 | ||
| M | North | 23 | 501–682 | 720–2 410 | – | 275–401 | – | – | |
| Centre | 130 | 439–672 | 317–2 250 | 298–2 120 | 259–384 | 205–329 | 250–346 | ||
| RJU | F | North | 19 | 538–860 | 1 300–6 500 | – | 407–595 | – | – |
| Centre | 91 | 517–933 | 814–7 291 | 748–6 197 | 347–655 | 282–512 | 286–447 | ||
| M | North | 24 | 480–860 | 650–5 000 | – | 355–550 | – | – | |
| Centre | 94 | 522–959 | 640–6 230 | 570–5 630 | 348–602 | 279–507 | 266–452 |
| Species | Sex | Area | n | TL | TW | gW (g) | DW (mm) | DL (mm) | CL (mm) |
|---|---|---|---|---|---|---|---|---|---|
| JAI | F | South | 16 | 329–488 | – | – | 228–336 | – | – |
| M | South | 22 | 313–492 | – | – | 215–310 | – | – | |
| RJC | F | North | 150 | 420–905 | 200–7 750 | – | 305–608 | – | – |
| Centre | 155 | 489–934 | 623–6 710 | 585–5 810 | 345–669 | 248–489 | 269–464 | ||
| South | 22 | 362–793 | 900–3 800 | – | 263–588 | – | – | ||
| M | North | 133 | 410–875 | 450–4 140 | – | 295–562 | – | – | |
| Centre | 80 | 458–870 | 540–3 870 | 480–3 435 | 321–565 | 226–428 | 260–443 | ||
| South | 30 | 315–850 | 1 000–3 200 | – | 212–547 | – | – | ||
| RJH | F | North | 68 | 380–975 | 400–8 520 | – | 297–715 | – | – |
| Centre | 148 | 376–1 061 | 304–10 680 | 283–9 160 | 268–771 | 205–588 | 230–514 | ||
| M | North | 67 | 389–1 005 | 400–8 700 | – | 284–723 | – | – | |
| Centre | 114 | 412–1 005 | 421–7 290 | 387–6 404 | 292–692 | 240–554 | 260–495 | ||
| RJM | F | North | 33 | 368–702 | 270–2 340 | – | 257–459 | – | – |
| Centre | 185 | 390–702 | 388–2 583 | 360–2 306 | 285–483 | 212–373 | 240–346 | ||
| South | 18 | 331–575 | – | – | 235–412 | – | – | ||
| M | North | 23 | 412–651 | 450–1 980 | – | 281–430 | – | – | |
| Centre | 97 | 408–612 | 455–1 575 | 420–1 450 | 292–412 | 209–319 | 226–336 | ||
| South | 20 | 366–522 | – | – | 253–354 | – | – | ||
| RJN | F | North | 23 | 497–718 | 700–2 460 | – | 270–409 | – | – |
| Centre | 224 | 448–691 | 460–2 135 | 435–1 975 | 249–399 | 215–340 | 250–353 | ||
| M | North | 23 | 501–682 | 720–2 410 | – | 275–401 | – | – | |
| Centre | 130 | 439–672 | 317–2 250 | 298–2 120 | 259–384 | 205–329 | 250–346 | ||
| RJU | F | North | 19 | 538–860 | 1 300–6 500 | – | 407–595 | – | – |
| Centre | 91 | 517–933 | 814–7 291 | 748–6 197 | 347–655 | 282–512 | 286–447 | ||
| M | North | 24 | 480–860 | 650–5 000 | – | 355–550 | – | – | |
| Centre | 94 | 522–959 | 640–6 230 | 570–5 630 | 348–602 | 279–507 | 266–452 |
TL, total length; TW, total weight; gW, gutted weight; DW, disc width; DL, disc length; CL, tail length; JAI, R. miraletus; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata.
Summary of the data available by species, sex, and area, provided as ranges for each morphometric measurement.
| Species | Sex | Area | n | TL | TW | gW (g) | DW (mm) | DL (mm) | CL (mm) |
|---|---|---|---|---|---|---|---|---|---|
| JAI | F | South | 16 | 329–488 | – | – | 228–336 | – | – |
| M | South | 22 | 313–492 | – | – | 215–310 | – | – | |
| RJC | F | North | 150 | 420–905 | 200–7 750 | – | 305–608 | – | – |
| Centre | 155 | 489–934 | 623–6 710 | 585–5 810 | 345–669 | 248–489 | 269–464 | ||
| South | 22 | 362–793 | 900–3 800 | – | 263–588 | – | – | ||
| M | North | 133 | 410–875 | 450–4 140 | – | 295–562 | – | – | |
| Centre | 80 | 458–870 | 540–3 870 | 480–3 435 | 321–565 | 226–428 | 260–443 | ||
| South | 30 | 315–850 | 1 000–3 200 | – | 212–547 | – | – | ||
| RJH | F | North | 68 | 380–975 | 400–8 520 | – | 297–715 | – | – |
| Centre | 148 | 376–1 061 | 304–10 680 | 283–9 160 | 268–771 | 205–588 | 230–514 | ||
| M | North | 67 | 389–1 005 | 400–8 700 | – | 284–723 | – | – | |
| Centre | 114 | 412–1 005 | 421–7 290 | 387–6 404 | 292–692 | 240–554 | 260–495 | ||
| RJM | F | North | 33 | 368–702 | 270–2 340 | – | 257–459 | – | – |
| Centre | 185 | 390–702 | 388–2 583 | 360–2 306 | 285–483 | 212–373 | 240–346 | ||
| South | 18 | 331–575 | – | – | 235–412 | – | – | ||
| M | North | 23 | 412–651 | 450–1 980 | – | 281–430 | – | – | |
| Centre | 97 | 408–612 | 455–1 575 | 420–1 450 | 292–412 | 209–319 | 226–336 | ||
| South | 20 | 366–522 | – | – | 253–354 | – | – | ||
| RJN | F | North | 23 | 497–718 | 700–2 460 | – | 270–409 | – | – |
| Centre | 224 | 448–691 | 460–2 135 | 435–1 975 | 249–399 | 215–340 | 250–353 | ||
| M | North | 23 | 501–682 | 720–2 410 | – | 275–401 | – | – | |
| Centre | 130 | 439–672 | 317–2 250 | 298–2 120 | 259–384 | 205–329 | 250–346 | ||
| RJU | F | North | 19 | 538–860 | 1 300–6 500 | – | 407–595 | – | – |
| Centre | 91 | 517–933 | 814–7 291 | 748–6 197 | 347–655 | 282–512 | 286–447 | ||
| M | North | 24 | 480–860 | 650–5 000 | – | 355–550 | – | – | |
| Centre | 94 | 522–959 | 640–6 230 | 570–5 630 | 348–602 | 279–507 | 266–452 |
| Species | Sex | Area | n | TL | TW | gW (g) | DW (mm) | DL (mm) | CL (mm) |
|---|---|---|---|---|---|---|---|---|---|
| JAI | F | South | 16 | 329–488 | – | – | 228–336 | – | – |
| M | South | 22 | 313–492 | – | – | 215–310 | – | – | |
| RJC | F | North | 150 | 420–905 | 200–7 750 | – | 305–608 | – | – |
| Centre | 155 | 489–934 | 623–6 710 | 585–5 810 | 345–669 | 248–489 | 269–464 | ||
| South | 22 | 362–793 | 900–3 800 | – | 263–588 | – | – | ||
| M | North | 133 | 410–875 | 450–4 140 | – | 295–562 | – | – | |
| Centre | 80 | 458–870 | 540–3 870 | 480–3 435 | 321–565 | 226–428 | 260–443 | ||
| South | 30 | 315–850 | 1 000–3 200 | – | 212–547 | – | – | ||
| RJH | F | North | 68 | 380–975 | 400–8 520 | – | 297–715 | – | – |
| Centre | 148 | 376–1 061 | 304–10 680 | 283–9 160 | 268–771 | 205–588 | 230–514 | ||
| M | North | 67 | 389–1 005 | 400–8 700 | – | 284–723 | – | – | |
| Centre | 114 | 412–1 005 | 421–7 290 | 387–6 404 | 292–692 | 240–554 | 260–495 | ||
| RJM | F | North | 33 | 368–702 | 270–2 340 | – | 257–459 | – | – |
| Centre | 185 | 390–702 | 388–2 583 | 360–2 306 | 285–483 | 212–373 | 240–346 | ||
| South | 18 | 331–575 | – | – | 235–412 | – | – | ||
| M | North | 23 | 412–651 | 450–1 980 | – | 281–430 | – | – | |
| Centre | 97 | 408–612 | 455–1 575 | 420–1 450 | 292–412 | 209–319 | 226–336 | ||
| South | 20 | 366–522 | – | – | 253–354 | – | – | ||
| RJN | F | North | 23 | 497–718 | 700–2 460 | – | 270–409 | – | – |
| Centre | 224 | 448–691 | 460–2 135 | 435–1 975 | 249–399 | 215–340 | 250–353 | ||
| M | North | 23 | 501–682 | 720–2 410 | – | 275–401 | – | – | |
| Centre | 130 | 439–672 | 317–2 250 | 298–2 120 | 259–384 | 205–329 | 250–346 | ||
| RJU | F | North | 19 | 538–860 | 1 300–6 500 | – | 407–595 | – | – |
| Centre | 91 | 517–933 | 814–7 291 | 748–6 197 | 347–655 | 282–512 | 286–447 | ||
| M | North | 24 | 480–860 | 650–5 000 | – | 355–550 | – | – | |
| Centre | 94 | 522–959 | 640–6 230 | 570–5 630 | 348–602 | 279–507 | 266–452 |
TL, total length; TW, total weight; gW, gutted weight; DW, disc width; DL, disc length; CL, tail length; JAI, R. miraletus; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata.
Morphometric variation within species
Apart from the variability observed between species, differences in the morphometric ratios (DW:TL, DL:TL, CL:TL and DL:DW) were also observed within species (Figure 3). Leucoraja naevus was clearly the species showing the most distinctive shape, relative to the other species. It has a narrower disc in relation to body length (lowest DW:TL) and the disc is longer than wide (higher DL:DW). Raja montagui showed the following morphometric similarities with the other three species: DL:TL and CL:TL with R. clavata; DL:DW with R. brachyura; and DW:TL with R. miraletus.
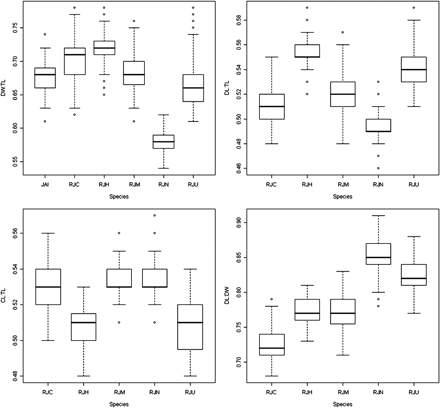
Boxplots showing species-specific variation in the morphometric ratios DW:TL, DL:TL, CL:TL, DL:DW. JAI, R. miraletus; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata. Boxplot statistical entries: maximum, third (75%) quartile, median, first (25%) quartile, and minimum, and the open circles represent outliers.
Nested models for the DW:TL ratio, considering the interactions of area, sex, and TL classes, were statistically significant for R. clavata, R. montagui, and R. undulata (p < 0.001). Nested models for the CL:TL ratio, considering the interaction between sex and TL classes, were statistically significant for all species except R. undulata (p < 0.01). The nested model for DL:DW, considering the interaction between sex and TL classes, was only significant for R. brachyura (p < 0.001). The size conversion factors estimated for the aforementioned nested models are presented in Table 2. Variations between area, sex, and TL class were observed, but there was no general trend.
Estimates of the nested models for the morphometric ratios (DW:TL, CL:TL, and DL:DW), by sex, area (N, north; C, centre; S, south), and TL class.
| Species | Ratio | R2 | TL class (mm) | Females | Males | ||||
|---|---|---|---|---|---|---|---|---|---|
| N | C | S | N | C | S | ||||
| RJC | DW:TL | 0.56 | 300 | 0.715 | 0.716 | 0.717 | 0.642 | 0.666 | 0.703 |
| 400 | 0.722 | 0.710 | 0.707 | 0.717 | 0.710 | 0.723 | |||
| 500 | 0.711 | 0.717 | 0.725 | 0.704 | 0.703 | 0.692 | |||
| 600 | 0.721 | 0.719 | 0.745 | 0.682 | 0.699 | 0.680 | |||
| 700 | 0.715 | 0.727 | 0.728 | 0.663 | 0.673 | 0.674 | |||
| 800 | 0.709 | 0.717 | – | – | – | 0.640 | |||
| CL:TL | 0.56 | 400 | 0.540 | 0.557 | |||||
| 500 | 0.536 | 0.457 | |||||||
| 600 | 0.527 | 0.537 | |||||||
| 700 | 0.515 | 0.540 | |||||||
| 800 | 0.522 | 0.540 | |||||||
| RJH | CL:TL | 0.54 | 400 | 0.515 | 0.510 | ||||
| 500 | 0.513 | 0.519 | |||||||
| 600 | 0.523 | 0.510 | |||||||
| 700 | 0.500 | 0.513 | |||||||
| 800 | 0.495 | 0.500 | |||||||
| 900 | – | – | |||||||
| 1 000 | 0.480 | – | |||||||
| DL:DW | 0.51 | 300 | 0.765 | 0.81 | |||||
| 400 | 0.753 | 0.765 | |||||||
| 500 | 0.752 | 0.757 | |||||||
| 600 | 0.76 | 0.766 | |||||||
| 700 | 0.771 | 0.776 | |||||||
| 800 | 0.782 | 0.787 | |||||||
| 900 | 0.78 | 0.788 | |||||||
| 1 000 | 0.772 | – | |||||||
| RJM | DW:TL | 0.51 | 300 | 0.7 | 0.74 | 0.714 | 0.644 | 0.662 | 0.701 |
| 400 | 0.6933 | 0.702 | 0.707 | 0.682 | 0.676 | 0.704 | |||
| 500 | 0.702 | 0.687 | 0.705 | 0.66 | 0.655 | 0.69 | |||
| 600 | 0.684 | 0.688 | – | – | – | – | |||
| 700 | 0.65 | 0.69 | – | – | – | – | |||
| CL:TL | 0.52 | 400 | 0.539 | 0.534 | |||||
| 500 | 0.533 | 0.546 | |||||||
| 600 | 0.523 | 0.551 | |||||||
| RJN | CL:TL | 0.51 | 400 | 0.55 | 0.55 | ||||
| 500 | 0.532 | 0.524 | |||||||
| 600 | 0.524 | 0.521 | |||||||
| RJU | DW:TL | 0.72 | 400 | 0.664 | 0.664 | – | 0.726 | 0.726 | – |
| 500 | 0.74 | 0.673 | – | 0.706 | 0.769 | – | |||
| 600 | 0.707 | 0.678 | – | 0.69 | 0.769 | – | |||
| 700 | 0.706 | 0.676 | – | 0.653 | 0.759 | – | |||
| 800 | – | 0.662 | – | 0.606 | 0.738 | – | |||
| Species | Ratio | R2 | TL class (mm) | Females | Males | ||||
|---|---|---|---|---|---|---|---|---|---|
| N | C | S | N | C | S | ||||
| RJC | DW:TL | 0.56 | 300 | 0.715 | 0.716 | 0.717 | 0.642 | 0.666 | 0.703 |
| 400 | 0.722 | 0.710 | 0.707 | 0.717 | 0.710 | 0.723 | |||
| 500 | 0.711 | 0.717 | 0.725 | 0.704 | 0.703 | 0.692 | |||
| 600 | 0.721 | 0.719 | 0.745 | 0.682 | 0.699 | 0.680 | |||
| 700 | 0.715 | 0.727 | 0.728 | 0.663 | 0.673 | 0.674 | |||
| 800 | 0.709 | 0.717 | – | – | – | 0.640 | |||
| CL:TL | 0.56 | 400 | 0.540 | 0.557 | |||||
| 500 | 0.536 | 0.457 | |||||||
| 600 | 0.527 | 0.537 | |||||||
| 700 | 0.515 | 0.540 | |||||||
| 800 | 0.522 | 0.540 | |||||||
| RJH | CL:TL | 0.54 | 400 | 0.515 | 0.510 | ||||
| 500 | 0.513 | 0.519 | |||||||
| 600 | 0.523 | 0.510 | |||||||
| 700 | 0.500 | 0.513 | |||||||
| 800 | 0.495 | 0.500 | |||||||
| 900 | – | – | |||||||
| 1 000 | 0.480 | – | |||||||
| DL:DW | 0.51 | 300 | 0.765 | 0.81 | |||||
| 400 | 0.753 | 0.765 | |||||||
| 500 | 0.752 | 0.757 | |||||||
| 600 | 0.76 | 0.766 | |||||||
| 700 | 0.771 | 0.776 | |||||||
| 800 | 0.782 | 0.787 | |||||||
| 900 | 0.78 | 0.788 | |||||||
| 1 000 | 0.772 | – | |||||||
| RJM | DW:TL | 0.51 | 300 | 0.7 | 0.74 | 0.714 | 0.644 | 0.662 | 0.701 |
| 400 | 0.6933 | 0.702 | 0.707 | 0.682 | 0.676 | 0.704 | |||
| 500 | 0.702 | 0.687 | 0.705 | 0.66 | 0.655 | 0.69 | |||
| 600 | 0.684 | 0.688 | – | – | – | – | |||
| 700 | 0.65 | 0.69 | – | – | – | – | |||
| CL:TL | 0.52 | 400 | 0.539 | 0.534 | |||||
| 500 | 0.533 | 0.546 | |||||||
| 600 | 0.523 | 0.551 | |||||||
| RJN | CL:TL | 0.51 | 400 | 0.55 | 0.55 | ||||
| 500 | 0.532 | 0.524 | |||||||
| 600 | 0.524 | 0.521 | |||||||
| RJU | DW:TL | 0.72 | 400 | 0.664 | 0.664 | – | 0.726 | 0.726 | – |
| 500 | 0.74 | 0.673 | – | 0.706 | 0.769 | – | |||
| 600 | 0.707 | 0.678 | – | 0.69 | 0.769 | – | |||
| 700 | 0.706 | 0.676 | – | 0.653 | 0.759 | – | |||
| 800 | – | 0.662 | – | 0.606 | 0.738 | – | |||
TL, total length; DW, disc width; DL, disc length; CL, tail length; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata.
Estimates of the nested models for the morphometric ratios (DW:TL, CL:TL, and DL:DW), by sex, area (N, north; C, centre; S, south), and TL class.
| Species | Ratio | R2 | TL class (mm) | Females | Males | ||||
|---|---|---|---|---|---|---|---|---|---|
| N | C | S | N | C | S | ||||
| RJC | DW:TL | 0.56 | 300 | 0.715 | 0.716 | 0.717 | 0.642 | 0.666 | 0.703 |
| 400 | 0.722 | 0.710 | 0.707 | 0.717 | 0.710 | 0.723 | |||
| 500 | 0.711 | 0.717 | 0.725 | 0.704 | 0.703 | 0.692 | |||
| 600 | 0.721 | 0.719 | 0.745 | 0.682 | 0.699 | 0.680 | |||
| 700 | 0.715 | 0.727 | 0.728 | 0.663 | 0.673 | 0.674 | |||
| 800 | 0.709 | 0.717 | – | – | – | 0.640 | |||
| CL:TL | 0.56 | 400 | 0.540 | 0.557 | |||||
| 500 | 0.536 | 0.457 | |||||||
| 600 | 0.527 | 0.537 | |||||||
| 700 | 0.515 | 0.540 | |||||||
| 800 | 0.522 | 0.540 | |||||||
| RJH | CL:TL | 0.54 | 400 | 0.515 | 0.510 | ||||
| 500 | 0.513 | 0.519 | |||||||
| 600 | 0.523 | 0.510 | |||||||
| 700 | 0.500 | 0.513 | |||||||
| 800 | 0.495 | 0.500 | |||||||
| 900 | – | – | |||||||
| 1 000 | 0.480 | – | |||||||
| DL:DW | 0.51 | 300 | 0.765 | 0.81 | |||||
| 400 | 0.753 | 0.765 | |||||||
| 500 | 0.752 | 0.757 | |||||||
| 600 | 0.76 | 0.766 | |||||||
| 700 | 0.771 | 0.776 | |||||||
| 800 | 0.782 | 0.787 | |||||||
| 900 | 0.78 | 0.788 | |||||||
| 1 000 | 0.772 | – | |||||||
| RJM | DW:TL | 0.51 | 300 | 0.7 | 0.74 | 0.714 | 0.644 | 0.662 | 0.701 |
| 400 | 0.6933 | 0.702 | 0.707 | 0.682 | 0.676 | 0.704 | |||
| 500 | 0.702 | 0.687 | 0.705 | 0.66 | 0.655 | 0.69 | |||
| 600 | 0.684 | 0.688 | – | – | – | – | |||
| 700 | 0.65 | 0.69 | – | – | – | – | |||
| CL:TL | 0.52 | 400 | 0.539 | 0.534 | |||||
| 500 | 0.533 | 0.546 | |||||||
| 600 | 0.523 | 0.551 | |||||||
| RJN | CL:TL | 0.51 | 400 | 0.55 | 0.55 | ||||
| 500 | 0.532 | 0.524 | |||||||
| 600 | 0.524 | 0.521 | |||||||
| RJU | DW:TL | 0.72 | 400 | 0.664 | 0.664 | – | 0.726 | 0.726 | – |
| 500 | 0.74 | 0.673 | – | 0.706 | 0.769 | – | |||
| 600 | 0.707 | 0.678 | – | 0.69 | 0.769 | – | |||
| 700 | 0.706 | 0.676 | – | 0.653 | 0.759 | – | |||
| 800 | – | 0.662 | – | 0.606 | 0.738 | – | |||
| Species | Ratio | R2 | TL class (mm) | Females | Males | ||||
|---|---|---|---|---|---|---|---|---|---|
| N | C | S | N | C | S | ||||
| RJC | DW:TL | 0.56 | 300 | 0.715 | 0.716 | 0.717 | 0.642 | 0.666 | 0.703 |
| 400 | 0.722 | 0.710 | 0.707 | 0.717 | 0.710 | 0.723 | |||
| 500 | 0.711 | 0.717 | 0.725 | 0.704 | 0.703 | 0.692 | |||
| 600 | 0.721 | 0.719 | 0.745 | 0.682 | 0.699 | 0.680 | |||
| 700 | 0.715 | 0.727 | 0.728 | 0.663 | 0.673 | 0.674 | |||
| 800 | 0.709 | 0.717 | – | – | – | 0.640 | |||
| CL:TL | 0.56 | 400 | 0.540 | 0.557 | |||||
| 500 | 0.536 | 0.457 | |||||||
| 600 | 0.527 | 0.537 | |||||||
| 700 | 0.515 | 0.540 | |||||||
| 800 | 0.522 | 0.540 | |||||||
| RJH | CL:TL | 0.54 | 400 | 0.515 | 0.510 | ||||
| 500 | 0.513 | 0.519 | |||||||
| 600 | 0.523 | 0.510 | |||||||
| 700 | 0.500 | 0.513 | |||||||
| 800 | 0.495 | 0.500 | |||||||
| 900 | – | – | |||||||
| 1 000 | 0.480 | – | |||||||
| DL:DW | 0.51 | 300 | 0.765 | 0.81 | |||||
| 400 | 0.753 | 0.765 | |||||||
| 500 | 0.752 | 0.757 | |||||||
| 600 | 0.76 | 0.766 | |||||||
| 700 | 0.771 | 0.776 | |||||||
| 800 | 0.782 | 0.787 | |||||||
| 900 | 0.78 | 0.788 | |||||||
| 1 000 | 0.772 | – | |||||||
| RJM | DW:TL | 0.51 | 300 | 0.7 | 0.74 | 0.714 | 0.644 | 0.662 | 0.701 |
| 400 | 0.6933 | 0.702 | 0.707 | 0.682 | 0.676 | 0.704 | |||
| 500 | 0.702 | 0.687 | 0.705 | 0.66 | 0.655 | 0.69 | |||
| 600 | 0.684 | 0.688 | – | – | – | – | |||
| 700 | 0.65 | 0.69 | – | – | – | – | |||
| CL:TL | 0.52 | 400 | 0.539 | 0.534 | |||||
| 500 | 0.533 | 0.546 | |||||||
| 600 | 0.523 | 0.551 | |||||||
| RJN | CL:TL | 0.51 | 400 | 0.55 | 0.55 | ||||
| 500 | 0.532 | 0.524 | |||||||
| 600 | 0.524 | 0.521 | |||||||
| RJU | DW:TL | 0.72 | 400 | 0.664 | 0.664 | – | 0.726 | 0.726 | – |
| 500 | 0.74 | 0.673 | – | 0.706 | 0.769 | – | |||
| 600 | 0.707 | 0.678 | – | 0.69 | 0.769 | – | |||
| 700 | 0.706 | 0.676 | – | 0.653 | 0.759 | – | |||
| 800 | – | 0.662 | – | 0.606 | 0.738 | – | |||
TL, total length; DW, disc width; DL, disc length; CL, tail length; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata.
For the remaining ratios (i.e. TL ∼ DW for R. miraletus, R. brachyura, and L. naevus, TL ∼ CL for R. undulata, TL ∼ DL for all species but R. miraletus, and DW ∼ DL for all species except R. miraletus and R. brachyura), there were linear relationships (Table 3). The estimates of the parameters for these relationships can be used as conversion factors between the various morphometric measurements. The coefficient of determination was high for all adjusted linear models (R2 > 0.85).
Parameter estimates of the linear models for the pairs of linear distances (DW ∼ TL, CL ∼ TL, DL ∼ TL, and DL ∼ DW), and the non-linear models, W ∼ a TLb and gW ∼ a TLb, with standard errors around the estimates presented in parenthesis.
| Relationship | Parameter | JAI | RJC | RJH | RJM | RJN | RJU |
|---|---|---|---|---|---|---|---|
| TL ∼ DW | a | −33.02 (27.18) | – | −17.78 (5.66) | – | 51.89 (7.46) | – |
| b | 1.60 (0.10) | – | 1.43 (0.01) | – | 1.57 (0.02) | – | |
| R2 | 0.88 | – | 0.98 | – | 0.93 | – | |
| TL ∼ CL | a | – | – | – | – | – | −34.66 (17.36) |
| b | – | – | – | – | – | 2.07 (0.05) | |
| R2 | – | – | – | – | – | 0.98 | |
| TL ∼ DL | a | – | 45.69 (8.83) | 32.62 (5.30) | 54.50 (7.86) | 66.37 (8.88) | 2.23 (12.77) |
| b | – | 1.83 (0.02) | 1.73 (0.01) | 1.72 (0.03) | 1.78 (0.03) | 1.84 (0.03) | |
| R2 | – | 0.97 | 0.99 | 0.94 | 0.91 | 0.96 | |
| DW ∼ DL | a | – | 10.12 (6.95) | – | 11.75 (5.96) | 17.98 (5.04) | 37.72 (7.32) |
| b | – | 1.36 (0.02) | – | 1.26 (0.02) | 1.11 (0.02) | 1.13 (0.02) | |
| R2 | – | 0.96 | – | 0.93 | 0.92 | 0.96 | |
| W ∼ a TLb | a | – | 5.20 × 10−6 (1.77 × 10−6) | 1.98 × 10−6 (5.07 × 10−7) | 3.44 × 10−7 (1.53 × 10−7) | 1.55 × 10−7 (7.45 × 10−8) | 1.92 × 10−5 (8.61 × 10−6) |
| b | – | 3.05 (0.05) | 3.20 (0.04) | 3.47 (0.07) | 3.58 (0.08) | 2.86 (0.07) | |
| gW ∼ a TLb | a | – | 5.71 × 10−6 (2.23 × 10−6) | 3.22 × 10−6 (8.12 × 10−7) | 8.03 × 10−7 (3.79 × 10−7) | 2.51 × 10−7 (1.27 × 10−7) | 4.91 × 10−6 (2.90 × 10−6) |
| b | – | 3.02 (0.06) | 3.11 (0.04) | 3.32 (0.07) | 3.49 (0.08) | 3.04 (0.09) |
| Relationship | Parameter | JAI | RJC | RJH | RJM | RJN | RJU |
|---|---|---|---|---|---|---|---|
| TL ∼ DW | a | −33.02 (27.18) | – | −17.78 (5.66) | – | 51.89 (7.46) | – |
| b | 1.60 (0.10) | – | 1.43 (0.01) | – | 1.57 (0.02) | – | |
| R2 | 0.88 | – | 0.98 | – | 0.93 | – | |
| TL ∼ CL | a | – | – | – | – | – | −34.66 (17.36) |
| b | – | – | – | – | – | 2.07 (0.05) | |
| R2 | – | – | – | – | – | 0.98 | |
| TL ∼ DL | a | – | 45.69 (8.83) | 32.62 (5.30) | 54.50 (7.86) | 66.37 (8.88) | 2.23 (12.77) |
| b | – | 1.83 (0.02) | 1.73 (0.01) | 1.72 (0.03) | 1.78 (0.03) | 1.84 (0.03) | |
| R2 | – | 0.97 | 0.99 | 0.94 | 0.91 | 0.96 | |
| DW ∼ DL | a | – | 10.12 (6.95) | – | 11.75 (5.96) | 17.98 (5.04) | 37.72 (7.32) |
| b | – | 1.36 (0.02) | – | 1.26 (0.02) | 1.11 (0.02) | 1.13 (0.02) | |
| R2 | – | 0.96 | – | 0.93 | 0.92 | 0.96 | |
| W ∼ a TLb | a | – | 5.20 × 10−6 (1.77 × 10−6) | 1.98 × 10−6 (5.07 × 10−7) | 3.44 × 10−7 (1.53 × 10−7) | 1.55 × 10−7 (7.45 × 10−8) | 1.92 × 10−5 (8.61 × 10−6) |
| b | – | 3.05 (0.05) | 3.20 (0.04) | 3.47 (0.07) | 3.58 (0.08) | 2.86 (0.07) | |
| gW ∼ a TLb | a | – | 5.71 × 10−6 (2.23 × 10−6) | 3.22 × 10−6 (8.12 × 10−7) | 8.03 × 10−7 (3.79 × 10−7) | 2.51 × 10−7 (1.27 × 10−7) | 4.91 × 10−6 (2.90 × 10−6) |
| b | – | 3.02 (0.06) | 3.11 (0.04) | 3.32 (0.07) | 3.49 (0.08) | 3.04 (0.09) |
TL, total length; DW, disc width; DL, disc length; CL, tail length; W, total weight; gW, gutted weight; JAI, R. miraletus; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata.
Parameter estimates of the linear models for the pairs of linear distances (DW ∼ TL, CL ∼ TL, DL ∼ TL, and DL ∼ DW), and the non-linear models, W ∼ a TLb and gW ∼ a TLb, with standard errors around the estimates presented in parenthesis.
| Relationship | Parameter | JAI | RJC | RJH | RJM | RJN | RJU |
|---|---|---|---|---|---|---|---|
| TL ∼ DW | a | −33.02 (27.18) | – | −17.78 (5.66) | – | 51.89 (7.46) | – |
| b | 1.60 (0.10) | – | 1.43 (0.01) | – | 1.57 (0.02) | – | |
| R2 | 0.88 | – | 0.98 | – | 0.93 | – | |
| TL ∼ CL | a | – | – | – | – | – | −34.66 (17.36) |
| b | – | – | – | – | – | 2.07 (0.05) | |
| R2 | – | – | – | – | – | 0.98 | |
| TL ∼ DL | a | – | 45.69 (8.83) | 32.62 (5.30) | 54.50 (7.86) | 66.37 (8.88) | 2.23 (12.77) |
| b | – | 1.83 (0.02) | 1.73 (0.01) | 1.72 (0.03) | 1.78 (0.03) | 1.84 (0.03) | |
| R2 | – | 0.97 | 0.99 | 0.94 | 0.91 | 0.96 | |
| DW ∼ DL | a | – | 10.12 (6.95) | – | 11.75 (5.96) | 17.98 (5.04) | 37.72 (7.32) |
| b | – | 1.36 (0.02) | – | 1.26 (0.02) | 1.11 (0.02) | 1.13 (0.02) | |
| R2 | – | 0.96 | – | 0.93 | 0.92 | 0.96 | |
| W ∼ a TLb | a | – | 5.20 × 10−6 (1.77 × 10−6) | 1.98 × 10−6 (5.07 × 10−7) | 3.44 × 10−7 (1.53 × 10−7) | 1.55 × 10−7 (7.45 × 10−8) | 1.92 × 10−5 (8.61 × 10−6) |
| b | – | 3.05 (0.05) | 3.20 (0.04) | 3.47 (0.07) | 3.58 (0.08) | 2.86 (0.07) | |
| gW ∼ a TLb | a | – | 5.71 × 10−6 (2.23 × 10−6) | 3.22 × 10−6 (8.12 × 10−7) | 8.03 × 10−7 (3.79 × 10−7) | 2.51 × 10−7 (1.27 × 10−7) | 4.91 × 10−6 (2.90 × 10−6) |
| b | – | 3.02 (0.06) | 3.11 (0.04) | 3.32 (0.07) | 3.49 (0.08) | 3.04 (0.09) |
| Relationship | Parameter | JAI | RJC | RJH | RJM | RJN | RJU |
|---|---|---|---|---|---|---|---|
| TL ∼ DW | a | −33.02 (27.18) | – | −17.78 (5.66) | – | 51.89 (7.46) | – |
| b | 1.60 (0.10) | – | 1.43 (0.01) | – | 1.57 (0.02) | – | |
| R2 | 0.88 | – | 0.98 | – | 0.93 | – | |
| TL ∼ CL | a | – | – | – | – | – | −34.66 (17.36) |
| b | – | – | – | – | – | 2.07 (0.05) | |
| R2 | – | – | – | – | – | 0.98 | |
| TL ∼ DL | a | – | 45.69 (8.83) | 32.62 (5.30) | 54.50 (7.86) | 66.37 (8.88) | 2.23 (12.77) |
| b | – | 1.83 (0.02) | 1.73 (0.01) | 1.72 (0.03) | 1.78 (0.03) | 1.84 (0.03) | |
| R2 | – | 0.97 | 0.99 | 0.94 | 0.91 | 0.96 | |
| DW ∼ DL | a | – | 10.12 (6.95) | – | 11.75 (5.96) | 17.98 (5.04) | 37.72 (7.32) |
| b | – | 1.36 (0.02) | – | 1.26 (0.02) | 1.11 (0.02) | 1.13 (0.02) | |
| R2 | – | 0.96 | – | 0.93 | 0.92 | 0.96 | |
| W ∼ a TLb | a | – | 5.20 × 10−6 (1.77 × 10−6) | 1.98 × 10−6 (5.07 × 10−7) | 3.44 × 10−7 (1.53 × 10−7) | 1.55 × 10−7 (7.45 × 10−8) | 1.92 × 10−5 (8.61 × 10−6) |
| b | – | 3.05 (0.05) | 3.20 (0.04) | 3.47 (0.07) | 3.58 (0.08) | 2.86 (0.07) | |
| gW ∼ a TLb | a | – | 5.71 × 10−6 (2.23 × 10−6) | 3.22 × 10−6 (8.12 × 10−7) | 8.03 × 10−7 (3.79 × 10−7) | 2.51 × 10−7 (1.27 × 10−7) | 4.91 × 10−6 (2.90 × 10−6) |
| b | – | 3.02 (0.06) | 3.11 (0.04) | 3.32 (0.07) | 3.49 (0.08) | 3.04 (0.09) |
TL, total length; DW, disc width; DL, disc length; CL, tail length; W, total weight; gW, gutted weight; JAI, R. miraletus; RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata.
According to the results of the likelihood-ratio test, the differences between the estimated parameters of the length ∼ weight models (TW ∼ TL and gW ∼ TL) adjusted by sex were not statistically different at a 95% confidence level, for all species. The parameter estimates of the non-linear models TW ∼ TL and gW ∼ TL are also presented in Table 3. The estimated allometric coefficients were between 2.86 (R. undulata) and 3.58 (L. naevus) for TW ∼ TL, and between 3.02 (R. clavata) and 3.49 (L. naevus) for gW ∼ TL.
Species discrimination using morphometric analysis
In a first FDA procedure, the species and sexes were entered as discriminant classes. The FDA results revealed no sexual dimorphism in the morphometric ratios for most of the species studied (Table 4). The misclassification error was 0.33. The species with the lowest misclassification between sexes was R. montagui (10% for females and 0% for males), followed by R. clavata (16% for males and 21% for females). Yet, R. montagui females were misclassified as R. clavata males in 20% and as R. brachyura females in 14%. The species with the greatest misclassification between sexes were R. brachyura and L. naevus, for both of which most males were identified as females, 92 and 77% accordingly.
Results of the FDA between five skate species and sexes (F, female; M, male).
| RJC (F) | RJC (M) | RJH (F) | RJH (M) | RJM (F) | RJM (M) | RJN (F) | RJN (M) | RJU (F) | RJU (M) | |
|---|---|---|---|---|---|---|---|---|---|---|
| RJC (F) | 78 | 21 | 2 | 0 | 0 | 5 | 0 | 0 | 0 | 0 |
| RJC (M) | 16 | 54 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 |
| RJH (F) | 6 | 4 | 91 | 92 | 14 | 10 | 0 | 0 | 0 | 0 |
| RJH (M) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| RJM (F) | 0 | 21 | 7 | 4 | 66 | 10 | 0 | 0 | 6 | 0 |
| RJM (M) | 0 | 0 | 0 | 0 | 0 | 75 | 0 | 0 | 6 | 0 |
| RJN (F) | 0 | 0 | 0 | 0 | 0 | 0 | 97 | 77 | 0 | 10 |
| RJN (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 23 | 0 | 5 |
| RJU (F) | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 61 | 43 |
| RJU (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 28 | 43 |
| RJC (F) | RJC (M) | RJH (F) | RJH (M) | RJM (F) | RJM (M) | RJN (F) | RJN (M) | RJU (F) | RJU (M) | |
|---|---|---|---|---|---|---|---|---|---|---|
| RJC (F) | 78 | 21 | 2 | 0 | 0 | 5 | 0 | 0 | 0 | 0 |
| RJC (M) | 16 | 54 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 |
| RJH (F) | 6 | 4 | 91 | 92 | 14 | 10 | 0 | 0 | 0 | 0 |
| RJH (M) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| RJM (F) | 0 | 21 | 7 | 4 | 66 | 10 | 0 | 0 | 6 | 0 |
| RJM (M) | 0 | 0 | 0 | 0 | 0 | 75 | 0 | 0 | 6 | 0 |
| RJN (F) | 0 | 0 | 0 | 0 | 0 | 0 | 97 | 77 | 0 | 10 |
| RJN (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 23 | 0 | 5 |
| RJU (F) | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 61 | 43 |
| RJU (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 28 | 43 |
Each column corresponds to the identified species and sexes and the rows correspond to the classification made by the FDA model. RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata. The cells of the matrix represent the percentage of the classification. Total misclassification error, 0.33.
Results of the FDA between five skate species and sexes (F, female; M, male).
| RJC (F) | RJC (M) | RJH (F) | RJH (M) | RJM (F) | RJM (M) | RJN (F) | RJN (M) | RJU (F) | RJU (M) | |
|---|---|---|---|---|---|---|---|---|---|---|
| RJC (F) | 78 | 21 | 2 | 0 | 0 | 5 | 0 | 0 | 0 | 0 |
| RJC (M) | 16 | 54 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 |
| RJH (F) | 6 | 4 | 91 | 92 | 14 | 10 | 0 | 0 | 0 | 0 |
| RJH (M) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| RJM (F) | 0 | 21 | 7 | 4 | 66 | 10 | 0 | 0 | 6 | 0 |
| RJM (M) | 0 | 0 | 0 | 0 | 0 | 75 | 0 | 0 | 6 | 0 |
| RJN (F) | 0 | 0 | 0 | 0 | 0 | 0 | 97 | 77 | 0 | 10 |
| RJN (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 23 | 0 | 5 |
| RJU (F) | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 61 | 43 |
| RJU (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 28 | 43 |
| RJC (F) | RJC (M) | RJH (F) | RJH (M) | RJM (F) | RJM (M) | RJN (F) | RJN (M) | RJU (F) | RJU (M) | |
|---|---|---|---|---|---|---|---|---|---|---|
| RJC (F) | 78 | 21 | 2 | 0 | 0 | 5 | 0 | 0 | 0 | 0 |
| RJC (M) | 16 | 54 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 |
| RJH (F) | 6 | 4 | 91 | 92 | 14 | 10 | 0 | 0 | 0 | 0 |
| RJH (M) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| RJM (F) | 0 | 21 | 7 | 4 | 66 | 10 | 0 | 0 | 6 | 0 |
| RJM (M) | 0 | 0 | 0 | 0 | 0 | 75 | 0 | 0 | 6 | 0 |
| RJN (F) | 0 | 0 | 0 | 0 | 0 | 0 | 97 | 77 | 0 | 10 |
| RJN (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 23 | 0 | 5 |
| RJU (F) | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 61 | 43 |
| RJU (M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 28 | 43 |
Each column corresponds to the identified species and sexes and the rows correspond to the classification made by the FDA model. RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata. The cells of the matrix represent the percentage of the classification. Total misclassification error, 0.33.
In the second procedure, only species were entered as discriminant classes. In this case, the morphometric ratios (DW:TL, DL:DW, and CL:DL) proved to be adequate for discriminating rajid species (Table 5). The misclassification error was 0.12. Leucoraja naevus was always discriminated from the other case-study species. The greatest proportion of misclassifications was between R. clavata (15%) and R. montagui (29%). The former was misclassified as R. montagui, and the latter as either R. clavata or R. brachyura.
Results of the FDA between five species of skate.
| RJC | RJH | RJM | RJN | RJU | |
|---|---|---|---|---|---|
| RJC | 85 | 1 | 18 | 0 | 0 |
| RJH | 3 | 91 | 11 | 0 | 5 |
| RJM | 12 | 6 | 71 | 0 | 5 |
| RJN | 0 | 0 | 0 | 100 | 8 |
| RJU | 0 | 1 | 0 | 0 | 82 |
| RJC | RJH | RJM | RJN | RJU | |
|---|---|---|---|---|---|
| RJC | 85 | 1 | 18 | 0 | 0 |
| RJH | 3 | 91 | 11 | 0 | 5 |
| RJM | 12 | 6 | 71 | 0 | 5 |
| RJN | 0 | 0 | 0 | 100 | 8 |
| RJU | 0 | 1 | 0 | 0 | 82 |
Each column corresponds to the species and the rows correspond to the classification made by the FDA model. RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata. The cells of the matrix represent the percentage of the classification. Total misclassification error, 0.12.
Results of the FDA between five species of skate.
| RJC | RJH | RJM | RJN | RJU | |
|---|---|---|---|---|---|
| RJC | 85 | 1 | 18 | 0 | 0 |
| RJH | 3 | 91 | 11 | 0 | 5 |
| RJM | 12 | 6 | 71 | 0 | 5 |
| RJN | 0 | 0 | 0 | 100 | 8 |
| RJU | 0 | 1 | 0 | 0 | 82 |
| RJC | RJH | RJM | RJN | RJU | |
|---|---|---|---|---|---|
| RJC | 85 | 1 | 18 | 0 | 0 |
| RJH | 3 | 91 | 11 | 0 | 5 |
| RJM | 12 | 6 | 71 | 0 | 5 |
| RJN | 0 | 0 | 0 | 100 | 8 |
| RJU | 0 | 1 | 0 | 0 | 82 |
Each column corresponds to the species and the rows correspond to the classification made by the FDA model. RJC, R. clavata; RJH, R. brachyura; RJM, R. montagui; RJN, L. naevus; RJU, R. undulata. The cells of the matrix represent the percentage of the classification. Total misclassification error, 0.12.
Discussion
Following the call made by the ICES WGEF (ICES, 2007), size conversion factors were estimated for the six skate species studied here. A clear definition of the measurements, including anterior and posterior reference points and how the distance between these two points was measured, allow future application of the results without additional bias (Francis, 2006). As proposed, the longest longitudinal axis (TL) was used in most size conversion factors, because it is considered to be the best index of size (Francis, 2006). In some species (e.g. R. clavata, R. montagui, and R. undulata) differences were recorded between sexes, areas, and size classes. These differences should be taken into consideration when applying the conversion factors to other subsets, and for that reason, the sex, area, sample size, and length range of dependent and independent variables were reported for each species. Only limited morphometric information has been presented by other authors, typically in association with reproduction and growth studies, with the TL ∼ W and TL ∼ DW relationships the most frequently reported (Du Buit, 1975; Nottage and Perkins, 1983; Ryland and Ajayi, 1984; Coelho and Erzini, 2002). Jardas (1975) was the only author to focus on the morphometry of R. clavata, but he did not provide conversion factors. Pallaoro et al. (2005) presented the W ∼ TL relationship for R. clavata and R. miraletus. Comparing the results from different studies, it is evident that, even for the same species, the conversion factors can vary between areas and sexes. The TL ∼ W relationship is merely indicative for a species, because weight varies with size class, and many external factors may also influence the total weight of an individual, and hence influence the TL ∼ W relationship, including maturation, spawning period, and food intake (Bagenal and Tesch, 1978). Moreover, the size range used in an analysis may also account for any differences observed between various studies. For that reason, the TL ∼ W relationships are only really applicable for the size range examined (Petrakis and Stergiou, 1995). Data on gW are limited, and such data could be usefully collected in future studies elsewhere in European waters. The gW could be more informative when comparing fish from different sampling areas exposed to different environmental conditions.
The results indicate that morphometric ratios may be considered a useful, simple, and reliable additional tool for helping with species discrimination. They can be applied successfully in cases where recurrent doubts remain on the separation of species, as is often the case between some specimens of R. montagui and R. brachyura, which can share a similar colour pattern, but do show distinct body morphometry, with a low percentage of misclassification between them. Also, if less specialized workers encounter problematic animals in the field, the collection of more information on the morphometry of such fish could serve as a useful tool for subsequently validating identification, without the need for examining more complex diagnostic characters (e.g. the number of rows of teeth, or clasper structure). Three morphometric ratios (DW:TL, DL:DW, and CL:DL) proved to be adequate for discriminating the six rajid species under study, according to FDA. The measurements proved to be 100% successful in identifying L. naevus, a species belonging to a different genus from the remaining five species, and therefore characterized by a defined morphotype, with narrow, long disc, and a long tail. Raja montagui and R. clavata provided the most cases of misclassification; both these species have a diamond-shaped disc that could be a reason for the observed misclassification given by FDA.
Acknowledgements
We thank Amador Lopes, Jorge Barra, Hélder Antunes, Augusto Pombal, José Lago, and Leonor Costa for collecting some of the data, and the reviewers, Jim Ellis and Joe Bizzarro, for providing useful comments and suggesting revisions to the English. The work was partially supported by the EU Data Collection Regulation/ PNAB. BS-P and TM were funded by grants from the Fundação para a Ciência e a Tecnologia, SFRH/BD/23777/2005 and SFRH/BD/29052/2006, respectively.

{kind=link}
{kind=link}
{kind=link}