





Abstract
The North Sea autumn-spawning herring (Clupea harengus) stock consists of a set of different spawning components. The dynamics of the entire stock have been well characterized, but although time-series of larval abundance indices are available for the individual components, study of the dynamics at the component level has historically been hampered by missing observations and high sampling noise. A simple state-space statistical model is developed that is robust to these problems, gives a good fit to the data, and proves capable of both handling and predicting missing observations well. Furthermore, the sum of the fitted abundance indices across all components proves an excellent proxy for the biomass of the total stock, even though the model utilizes information at the individual-component level. The Orkney–Shetland component appears to have recovered faster from historic depletion events than the other components, whereas the Downs component has been the slowest. These differences give rise to changes in stock composition, which are shown to vary widely within a relatively short time. The modelling framework provides a valuable tool for studying and monitoring the dynamics of the individual components of the North Sea herring stock.Payne, M. R. 2010. Mind the gaps: a state-space model for analysing the dynamics of North Sea herring spawning components. – ICES Journal of Marine Science, 67: 1939–1947.
Introduction
The North Sea autumn-spawning herring (Clupea harengus Linnaeus 1758) stock is generally understood as representing a complex of multiple spawning components (Cushing, 1955; Harden Jones, 1968; Iles and Sinclair, 1982; Heath et al., 1997). Most authors distinguish four major components (Figure 1), each defined by distinct spawning times and sites (Iles and Sinclair, 1982; Corten, 1986; Heath et al., 1997). The Orkney–Shetland component spawns in August/September between the islands that give it its name; the Buchan component to the east of Scotland in September/October; the Banks component off the English coast around the same time; and the Downs component in the English Channel during December. Although the different components mix outside the spawning season and are exploited together, each component is thought to have a high degree of population integrity (Iles and Sinclair, 1982) and, therefore, could be expected to have relatively unique population dynamics.
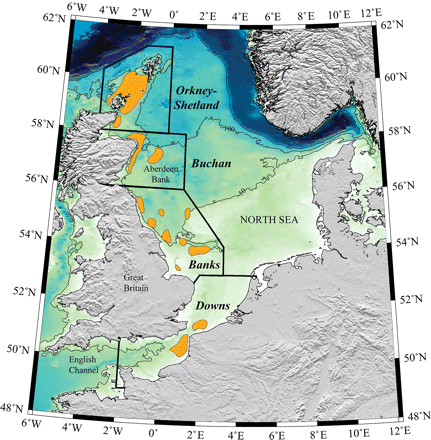
Map of the North Sea with recognized spawning grounds (Iles and Sinclair, 1982; Corten, 1986; Heath et al., 1997; Nash et al., 2009). Approximate boundaries of the areas covered by the IHLS are marked with bold lines. Depth contours are illustrated for 50, 100, and 200 m.
At the overall stock level, the biomass has varied widely during the past 50 years, collapsing from a late-1940s high of more than 5 million tonnes to a low of 50 000 t in 1976 (ICES 1998, 2009; Dickey-Collas et al., 2010), before recovering to peaks of 1.2 million tonnes in 1989 and 1.8 million tonnes in 2004 (Simmonds, 2007). In terms of individual spawning components, the stock collapsed progressively from the south to the north, with the southern Downs component collapsing as early as the 1950s and the northern Orkney–Shetland component becoming dominant by the 1970s (Burd, 1985; Corten, 1986; Cushing, 1992; Heath et al., 1997). The Downs component has only begun to recover to a substantial degree in recent years (ICES, 2009). Such knowledge, however, is qualitative in nature: a precise, quantitative understanding of the dynamics of the individual components remains lacking.
The annual international herring larval survey (IHLS) is designed to provide estimates of the larval abundance associated with the individual components, which in turn can be interpreted as a proxy for adult biomass (Postuma and Zijlstra, 1974; Heath, 1993). This survey is considered to have been consistent since 1972 and, therefore, potentially represents an excellent source of information covering both the period of stock collapse and recent recovery (Heath, 1993). The survey design is centred upon the concept of “sampling units”, the surveying of the spawning grounds of a given component during a specific time of the year (typically a half-month period). The full spawning period of each component is covered by multiple sampling units (typically between two and four), thereby generating several independent estimates of larval abundance. However, bad weather, restrictions on research-vessel availability, and more recently, cuts in funding for this survey have meant that the full survey has been carried out rarely according to the design: since 1990, an average of only 60% of the sampling units were covered each year. The resulting time-series is therefore plagued by large gaps, making comparisons between components difficult.
Several authors have tried to circumvent these problems and to develop an index robust to missing values. The multiplicative larval abundance index, MLAI (Gröger et al., 2001 and references therein), is based on the application of a multifactor ANOVA, where the year effects are used as the index, and it is currently used as an index of the total spawning-stock biomass (SSB) in the assessment of the entire North Sea stock (ICES, 2009; Simmonds, 2009). However, an implicit assumption in this approach is that the relative proportions between spawning components are fixed in time: by design, the MLAI cannot give information about changes in stock composition. Schmidt et al. (2009) examined the relative dynamics of the different components by selecting the most consistently sampled sampling unit for the Orkney–Shetland and Buchan components and taking the arithmetic mean across all units for the other components. However, such an approach omits a substantial fraction of the available data, is vulnerable to systematic differences among sampling units, and is susceptible to missing observations.
I present a simple statistical model to estimate the abundances of each spawning component in a manner robust to missing observations, to systematic differences among sampling units, and to statistical sampling noise. The model is based around the estimation of an index for the larval abundance (and hence adult biomass) for each particular spawning component and year. The key aspect of this “spawning-component abundance index” (SCAI) is that, by accounting for the issues mentioned earlier, it is readily comparable between both spawning components and years. The SCAI can therefore be used to examine the dynamics of the individual spawning components in relation to the total spawning stock.
Methods
Data sources
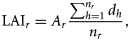
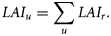
Characteristics of the individual sampling units used in the IHLS (ICES, 2006).
| Spawning component | Sampling-unit code | Sampling period | Coverage (%) |
|---|---|---|---|
| Orkney–Shetland | OS1 | 1–15 September | 62 |
| OS2 | 16–30 September | 100 | |
| Buchan | B1 | 1–15 September | 49 |
| B2 | 16–30 September | 89 | |
| Banks (Central NS) | CNS1 | 1–15 September | 57 |
| CNS2 | 16–30 September | 89 | |
| CNS3 | 1–15 October | 70 | |
| CNS4a | 16–31 October | 41 | |
| Downs (Southern NS) | SNS1 | 15–31 December | 89 |
| SNS2 | 1–15 January | 92 | |
| SNS3 | 16–31 January | 70 |
| Spawning component | Sampling-unit code | Sampling period | Coverage (%) |
|---|---|---|---|
| Orkney–Shetland | OS1 | 1–15 September | 62 |
| OS2 | 16–30 September | 100 | |
| Buchan | B1 | 1–15 September | 49 |
| B2 | 16–30 September | 89 | |
| Banks (Central NS) | CNS1 | 1–15 September | 57 |
| CNS2 | 16–30 September | 89 | |
| CNS3 | 1–15 October | 70 | |
| CNS4a | 16–31 October | 41 | |
| Downs (Southern NS) | SNS1 | 15–31 December | 89 |
| SNS2 | 1–15 January | 92 | |
| SNS3 | 16–31 January | 70 |
Coverage is calculated as the percentage of years (1972–2008) for which a LAI is available.
aThe CNS4 sampling unit was last surveyed in 1989: coverage up to that point was 83%.
Characteristics of the individual sampling units used in the IHLS (ICES, 2006).
| Spawning component | Sampling-unit code | Sampling period | Coverage (%) |
|---|---|---|---|
| Orkney–Shetland | OS1 | 1–15 September | 62 |
| OS2 | 16–30 September | 100 | |
| Buchan | B1 | 1–15 September | 49 |
| B2 | 16–30 September | 89 | |
| Banks (Central NS) | CNS1 | 1–15 September | 57 |
| CNS2 | 16–30 September | 89 | |
| CNS3 | 1–15 October | 70 | |
| CNS4a | 16–31 October | 41 | |
| Downs (Southern NS) | SNS1 | 15–31 December | 89 |
| SNS2 | 1–15 January | 92 | |
| SNS3 | 16–31 January | 70 |
| Spawning component | Sampling-unit code | Sampling period | Coverage (%) |
|---|---|---|---|
| Orkney–Shetland | OS1 | 1–15 September | 62 |
| OS2 | 16–30 September | 100 | |
| Buchan | B1 | 1–15 September | 49 |
| B2 | 16–30 September | 89 | |
| Banks (Central NS) | CNS1 | 1–15 September | 57 |
| CNS2 | 16–30 September | 89 | |
| CNS3 | 1–15 October | 70 | |
| CNS4a | 16–31 October | 41 | |
| Downs (Southern NS) | SNS1 | 15–31 December | 89 |
| SNS2 | 1–15 January | 92 | |
| SNS3 | 16–31 January | 70 |
Coverage is calculated as the percentage of years (1972–2008) for which a LAI is available.
aThe CNS4 sampling unit was last surveyed in 1989: coverage up to that point was 83%.
This analysis is based on the LAI estimated for each sampling unit and reported annually in the reports of the ICES Working Group for the Assessment of Herring South of 62°N (e.g. ICES, 2009).
Model assumptions and structure
The model is structured around the following assumptions: The resulting state-space model is conceptually similar to a delay-difference model, such as the Schaefer production model (Hilborn and Walters, 1992) or the biomass random-effects model (Trenkel, 2008), where the dynamics of the entire stock (such as recruitment, mortality, and growth) are treated as unobserved processes, and observations provide (uncertain) information. Although it might be possible to extend the SCAI model to consider these processes, with only a single data source available (the LAIs), the model would be susceptible to overparametrization. The relationship between successive population sizes [Equation (3)] has therefore been modelled for a single process, representing the net sum of recruitment, natural mortality, fishing mortality, and growth.
Assumption I: The individual spawning components display a high degree of population integrity, i.e. their population dynamics are predominantly controlled by the recruitment and mortality rates they experience and not by exchanges (immigration or emigration) with other components (Iles and Sinclair, 1982; Kell et al., 2009). It is therefore possible—and indeed necessary—to consider the dynamics of each component independently. The subsequent model is therefore described for a single component, but applied sequentially to estimate the SCAI for each component in turn.
- Assumption II: The size of a component in a given year is influenced by the size of the component in the previous year. For herring stocks, the SSB usually consists of multiple year classes and, therefore, a proportion of the adult population is carried over from year to year (ICES, 2009). Noting assumption I, assumption II is therefore applied at the component level as well, by writing the following relationship between the spawning component biomasses (SCB) in years y and y + 1:where the relative changes in SCB from year to year are assumed to be lognormally distributed with location parameter μSCB and scale parameter σSCB. Under the assumption that the larval abundance is proportional to the local spawning biomass of herring (Postuma and Zijlstra, 1974; Heath, 1993), Equation (3) can be recast for SCAI:(3)
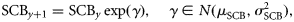 This result is equivalent to representing log(SCAI) as a normally distributed random walk with a mean step of μSCAI and variance(4)
This result is equivalent to representing log(SCAI) as a normally distributed random walk with a mean step of μSCAI and variance(4)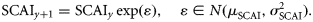
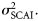
- Assumption III: The distribution of spawning intensity within each spawning component over time is relatively constant among years. The sampling units in the IHLS for a given component generally have comparable spatial coverage: the main distinction between them therefore is the time when they are done. Given that the intensity of spawning varies throughout the year (i.e. there is a distinct spawning season), an unknown proportion, pu, of the total amount of spawning (between 0 and 1) will be observed by a given sampling unit. It is therefore possible to relate the LAI for a given component, sampling unit and year, LAIu,y, to the SCAI for that component and year, SCAIy via the proportion pu:where the exponential term allows for a lognormally distributed error in the LAI observations. The precision of the survey,(5)
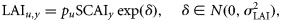
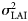 , is assumed the same across all years and all sampling units within a spawning component.
, is assumed the same across all years and all sampling units within a spawning component. - Assumption IV: The IHLS sampling units cover the entire spawning season and the spatial extent of spawning for each component. Therefore, there is no spawning outside of the area and the period covered in the survey design. In terms of Equation (5), the sum of the proportions over the k sampling units for any spawning component is therefore unity:This assumption refers explicitly to the design of the survey, rather than its execution: if the survey is executed fully and all sampling units are covered, the sum of the resultant LAIs is assumed to be an estimator of the component stock size. Missing sampling units in a given year are therefore not problematic, as long as the design covers the full spawning period of each component (and all sampling units are done at some point).(6)
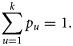
Parameter estimation
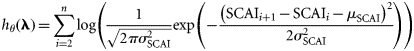
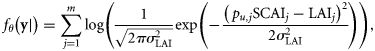
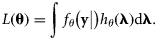
Stock assessment time-series
For comparison with the output of the model, the time-series of SSB of North Sea herring was taken from the most recent analytical assessment (ICES, 2009). However, the two datasets are not completely independent of each other, because the assessment includes information from the IHLS. Therefore, a second time-series of SSB estimates, excluding all IHLS information, but keeping the other input (i.e. catch data, trawl surveys, acoustic surveys, and other larval surveys), was produced by modifying the script used to do the assessment (revision 291, available from the HAWG assessment repository, http://hawg.googlecode.com).
Results
The model proved not to be overparametrized and fitted the observations for each spawning component well (Figure 2, Table 2), with the LAI observations distributed evenly around the fitted SCAI and the majority of points lying within the 95% confidence interval. The fitted model is also consistent with some of the underlying assumptions. The hypothesis of normally distributed interannual changes in the log(SCAI) index cannot be rejected (Kolmogorov–Smirnov tests: Orkney–Shetland p = 0.08, Buchan p = 0.45, Banks p = 0.15, Downs p = 0.24), consistent with Equation (4). In line with Equation (5), the null hypothesis of normally distributed residuals is not rejected by the Kolmogorov–Smirnov test at the 95% level for any of the 11 sampling units, and only for two using the Shapiro–Wilk test for normality.
SCAI with the s.e. of the log(SCAI) for each spawning component (cf. Figure 2) and year [95% confidence interval can be obtained from exp(log(SCAI) ± 1.96 s.e.)].
| SCAI | s.e. of Log(SCAI) | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | OrkShe | Buchan | Banks | Downs | OrkShe | Buchan | Banks | Downs |
| 1972 | 2 563 | 29 | 818 | 13 | 0.62 | 0.87 | 0.52 | 0.57 |
| 1973 | 1 890 | 17 | 1445 | 11 | 0.50 | 0.71 | 0.50 | 0.53 |
| 1974 | 1 151 | 183 | 902 | 11 | 0.46 | 0.71 | 0.52 | 0.51 |
| 1975 | 699 | 160 | 501 | 8 | 0.52 | 0.81 | 0.50 | 0.53 |
| 1976 | 717 | 24 | 434 | 11 | 0.51 | 0.99 | 0.49 | 0.58 |
| 1977 | 985 | 109 | 507 | 15 | 0.47 | 0.63 | 0.45 | 0.62 |
| 1978 | 1 323 | 216 | 582 | 41 | 0.46 | 0.86 | 0.44 | 0.48 |
| 1979 | 2 370 | 83 | 662 | 162 | 0.48 | 0.63 | 0.44 | 0.46 |
| 1980 | 2 671 | 18 | 540 | 324 | 0.46 | 0.70 | 0.46 | 0.43 |
| 1981 | 2 683 | 27 | 764 | 563 | 0.46 | 0.72 | 0.47 | 0.47 |
| 1982 | 3 132 | 466 | 913 | 592 | 0.46 | 0.63 | 0.45 | 0.41 |
| 1983 | 3 537 | 2 567 | 1 460 | 489 | 0.46 | 0.65 | 0.47 | 0.39 |
| 1984 | 4 420 | 4 038 | 3 496 | 579 | 0.46 | 0.62 | 0.46 | 0.39 |
| 1985 | 6 029 | 4 153 | 4 583 | 747 | 0.46 | 0.61 | 0.45 | 0.39 |
| 1986 | 6 543 | 2 783 | 4 401 | 683 | 0.46 | 0.62 | 0.44 | 0.40 |
| 1987 | 8 079 | 3 500 | 5 817 | 1 047 | 0.46 | 0.62 | 0.45 | 0.39 |
| 1988 | 10 349 | 8 753 | 6 834 | 1 427 | 0.50 | 0.64 | 0.46 | 0.39 |
| 1989 | 10 086 | 5 323 | 4 413 | 2 608 | 0.53 | 0.61 | 0.44 | 0.41 |
| 1990 | 7 399 | 6 126 | 4 798 | 3 061 | 0.55 | 0.62 | 0.50 | 0.45 |
| 1991 | 3 801 | 5 077 | 2 956 | 2 918 | 0.47 | 0.80 | 0.49 | 0.45 |
| 1992 | 2 153 | 2 443 | 811 | 2 028 | 0.51 | 0.81 | 0.54 | 0.44 |
| 1993 | 1 281 | 923 | 765 | 1 938 | 0.68 | 0.88 | 0.55 | 0.43 |
| 1994 | 1 363 | 752 | 705 | 1 409 | 0.63 | 1.32 | 0.57 | 0.43 |
| 1995 | 3 108 | 612 | 585 | 1 058 | 0.54 | 1.32 | 0.65 | 0.44 |
| 1996 | 4 049 | 499 | 678 | 1 763 | 0.56 | 0.86 | 0.66 | 0.39 |
| 1997 | 6 402 | 272 | 635 | 2 802 | 0.58 | 0.89 | 0.72 | 0.41 |
| 1998 | 9 133 | 1 290 | 595 | 2 494 | 0.61 | 0.91 | 0.62 | 0.39 |
| 1999 | 10 182 | 561 | 823 | 2 586 | 0.59 | 0.79 | 0.60 | 0.39 |
| 2000 | 11 305 | 199 | 1 508 | 2 881 | 0.59 | 0.71 | 0.63 | 0.39 |
| 2001 | 13 608 | 637 | 3 189 | 4 233 | 0.61 | 0.79 | 0.64 | 0.39 |
| 2002 | 13 035 | 2 551 | 6 326 | 4 515 | 0.59 | 0.82 | 0.66 | 0.40 |
| 2003 | 11 585 | 5 763 | 9 794 | 8 157 | 0.57 | 0.84 | 0.63 | 0.40 |
| 2004 | 11 392 | 7 816 | 9 705 | 10 067 | 0.57 | 0.85 | 0.66 | 0.40 |
| 2005 | 9 843 | 4 223 | 8 684 | 9 523 | 0.55 | 0.81 | 0.67 | 0.39 |
| 2006 | 8 841 | 1 925 | 6 763 | 11 837 | 0.51 | 0.84 | 0.69 | 0.39 |
| 2007 | 8 517 | 2 974 | 5 579 | 12 851 | 0.56 | 0.82 | 0.75 | 0.40 |
| 2008 | 9 872 | 2 274 | 8 462 | 15 119 | 0.59 | 0.92 | 0.85 | 0.46 |
| SCAI | s.e. of Log(SCAI) | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | OrkShe | Buchan | Banks | Downs | OrkShe | Buchan | Banks | Downs |
| 1972 | 2 563 | 29 | 818 | 13 | 0.62 | 0.87 | 0.52 | 0.57 |
| 1973 | 1 890 | 17 | 1445 | 11 | 0.50 | 0.71 | 0.50 | 0.53 |
| 1974 | 1 151 | 183 | 902 | 11 | 0.46 | 0.71 | 0.52 | 0.51 |
| 1975 | 699 | 160 | 501 | 8 | 0.52 | 0.81 | 0.50 | 0.53 |
| 1976 | 717 | 24 | 434 | 11 | 0.51 | 0.99 | 0.49 | 0.58 |
| 1977 | 985 | 109 | 507 | 15 | 0.47 | 0.63 | 0.45 | 0.62 |
| 1978 | 1 323 | 216 | 582 | 41 | 0.46 | 0.86 | 0.44 | 0.48 |
| 1979 | 2 370 | 83 | 662 | 162 | 0.48 | 0.63 | 0.44 | 0.46 |
| 1980 | 2 671 | 18 | 540 | 324 | 0.46 | 0.70 | 0.46 | 0.43 |
| 1981 | 2 683 | 27 | 764 | 563 | 0.46 | 0.72 | 0.47 | 0.47 |
| 1982 | 3 132 | 466 | 913 | 592 | 0.46 | 0.63 | 0.45 | 0.41 |
| 1983 | 3 537 | 2 567 | 1 460 | 489 | 0.46 | 0.65 | 0.47 | 0.39 |
| 1984 | 4 420 | 4 038 | 3 496 | 579 | 0.46 | 0.62 | 0.46 | 0.39 |
| 1985 | 6 029 | 4 153 | 4 583 | 747 | 0.46 | 0.61 | 0.45 | 0.39 |
| 1986 | 6 543 | 2 783 | 4 401 | 683 | 0.46 | 0.62 | 0.44 | 0.40 |
| 1987 | 8 079 | 3 500 | 5 817 | 1 047 | 0.46 | 0.62 | 0.45 | 0.39 |
| 1988 | 10 349 | 8 753 | 6 834 | 1 427 | 0.50 | 0.64 | 0.46 | 0.39 |
| 1989 | 10 086 | 5 323 | 4 413 | 2 608 | 0.53 | 0.61 | 0.44 | 0.41 |
| 1990 | 7 399 | 6 126 | 4 798 | 3 061 | 0.55 | 0.62 | 0.50 | 0.45 |
| 1991 | 3 801 | 5 077 | 2 956 | 2 918 | 0.47 | 0.80 | 0.49 | 0.45 |
| 1992 | 2 153 | 2 443 | 811 | 2 028 | 0.51 | 0.81 | 0.54 | 0.44 |
| 1993 | 1 281 | 923 | 765 | 1 938 | 0.68 | 0.88 | 0.55 | 0.43 |
| 1994 | 1 363 | 752 | 705 | 1 409 | 0.63 | 1.32 | 0.57 | 0.43 |
| 1995 | 3 108 | 612 | 585 | 1 058 | 0.54 | 1.32 | 0.65 | 0.44 |
| 1996 | 4 049 | 499 | 678 | 1 763 | 0.56 | 0.86 | 0.66 | 0.39 |
| 1997 | 6 402 | 272 | 635 | 2 802 | 0.58 | 0.89 | 0.72 | 0.41 |
| 1998 | 9 133 | 1 290 | 595 | 2 494 | 0.61 | 0.91 | 0.62 | 0.39 |
| 1999 | 10 182 | 561 | 823 | 2 586 | 0.59 | 0.79 | 0.60 | 0.39 |
| 2000 | 11 305 | 199 | 1 508 | 2 881 | 0.59 | 0.71 | 0.63 | 0.39 |
| 2001 | 13 608 | 637 | 3 189 | 4 233 | 0.61 | 0.79 | 0.64 | 0.39 |
| 2002 | 13 035 | 2 551 | 6 326 | 4 515 | 0.59 | 0.82 | 0.66 | 0.40 |
| 2003 | 11 585 | 5 763 | 9 794 | 8 157 | 0.57 | 0.84 | 0.63 | 0.40 |
| 2004 | 11 392 | 7 816 | 9 705 | 10 067 | 0.57 | 0.85 | 0.66 | 0.40 |
| 2005 | 9 843 | 4 223 | 8 684 | 9 523 | 0.55 | 0.81 | 0.67 | 0.39 |
| 2006 | 8 841 | 1 925 | 6 763 | 11 837 | 0.51 | 0.84 | 0.69 | 0.39 |
| 2007 | 8 517 | 2 974 | 5 579 | 12 851 | 0.56 | 0.82 | 0.75 | 0.40 |
| 2008 | 9 872 | 2 274 | 8 462 | 15 119 | 0.59 | 0.92 | 0.85 | 0.46 |
SCAI with the s.e. of the log(SCAI) for each spawning component (cf. Figure 2) and year [95% confidence interval can be obtained from exp(log(SCAI) ± 1.96 s.e.)].
| SCAI | s.e. of Log(SCAI) | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | OrkShe | Buchan | Banks | Downs | OrkShe | Buchan | Banks | Downs |
| 1972 | 2 563 | 29 | 818 | 13 | 0.62 | 0.87 | 0.52 | 0.57 |
| 1973 | 1 890 | 17 | 1445 | 11 | 0.50 | 0.71 | 0.50 | 0.53 |
| 1974 | 1 151 | 183 | 902 | 11 | 0.46 | 0.71 | 0.52 | 0.51 |
| 1975 | 699 | 160 | 501 | 8 | 0.52 | 0.81 | 0.50 | 0.53 |
| 1976 | 717 | 24 | 434 | 11 | 0.51 | 0.99 | 0.49 | 0.58 |
| 1977 | 985 | 109 | 507 | 15 | 0.47 | 0.63 | 0.45 | 0.62 |
| 1978 | 1 323 | 216 | 582 | 41 | 0.46 | 0.86 | 0.44 | 0.48 |
| 1979 | 2 370 | 83 | 662 | 162 | 0.48 | 0.63 | 0.44 | 0.46 |
| 1980 | 2 671 | 18 | 540 | 324 | 0.46 | 0.70 | 0.46 | 0.43 |
| 1981 | 2 683 | 27 | 764 | 563 | 0.46 | 0.72 | 0.47 | 0.47 |
| 1982 | 3 132 | 466 | 913 | 592 | 0.46 | 0.63 | 0.45 | 0.41 |
| 1983 | 3 537 | 2 567 | 1 460 | 489 | 0.46 | 0.65 | 0.47 | 0.39 |
| 1984 | 4 420 | 4 038 | 3 496 | 579 | 0.46 | 0.62 | 0.46 | 0.39 |
| 1985 | 6 029 | 4 153 | 4 583 | 747 | 0.46 | 0.61 | 0.45 | 0.39 |
| 1986 | 6 543 | 2 783 | 4 401 | 683 | 0.46 | 0.62 | 0.44 | 0.40 |
| 1987 | 8 079 | 3 500 | 5 817 | 1 047 | 0.46 | 0.62 | 0.45 | 0.39 |
| 1988 | 10 349 | 8 753 | 6 834 | 1 427 | 0.50 | 0.64 | 0.46 | 0.39 |
| 1989 | 10 086 | 5 323 | 4 413 | 2 608 | 0.53 | 0.61 | 0.44 | 0.41 |
| 1990 | 7 399 | 6 126 | 4 798 | 3 061 | 0.55 | 0.62 | 0.50 | 0.45 |
| 1991 | 3 801 | 5 077 | 2 956 | 2 918 | 0.47 | 0.80 | 0.49 | 0.45 |
| 1992 | 2 153 | 2 443 | 811 | 2 028 | 0.51 | 0.81 | 0.54 | 0.44 |
| 1993 | 1 281 | 923 | 765 | 1 938 | 0.68 | 0.88 | 0.55 | 0.43 |
| 1994 | 1 363 | 752 | 705 | 1 409 | 0.63 | 1.32 | 0.57 | 0.43 |
| 1995 | 3 108 | 612 | 585 | 1 058 | 0.54 | 1.32 | 0.65 | 0.44 |
| 1996 | 4 049 | 499 | 678 | 1 763 | 0.56 | 0.86 | 0.66 | 0.39 |
| 1997 | 6 402 | 272 | 635 | 2 802 | 0.58 | 0.89 | 0.72 | 0.41 |
| 1998 | 9 133 | 1 290 | 595 | 2 494 | 0.61 | 0.91 | 0.62 | 0.39 |
| 1999 | 10 182 | 561 | 823 | 2 586 | 0.59 | 0.79 | 0.60 | 0.39 |
| 2000 | 11 305 | 199 | 1 508 | 2 881 | 0.59 | 0.71 | 0.63 | 0.39 |
| 2001 | 13 608 | 637 | 3 189 | 4 233 | 0.61 | 0.79 | 0.64 | 0.39 |
| 2002 | 13 035 | 2 551 | 6 326 | 4 515 | 0.59 | 0.82 | 0.66 | 0.40 |
| 2003 | 11 585 | 5 763 | 9 794 | 8 157 | 0.57 | 0.84 | 0.63 | 0.40 |
| 2004 | 11 392 | 7 816 | 9 705 | 10 067 | 0.57 | 0.85 | 0.66 | 0.40 |
| 2005 | 9 843 | 4 223 | 8 684 | 9 523 | 0.55 | 0.81 | 0.67 | 0.39 |
| 2006 | 8 841 | 1 925 | 6 763 | 11 837 | 0.51 | 0.84 | 0.69 | 0.39 |
| 2007 | 8 517 | 2 974 | 5 579 | 12 851 | 0.56 | 0.82 | 0.75 | 0.40 |
| 2008 | 9 872 | 2 274 | 8 462 | 15 119 | 0.59 | 0.92 | 0.85 | 0.46 |
| SCAI | s.e. of Log(SCAI) | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | OrkShe | Buchan | Banks | Downs | OrkShe | Buchan | Banks | Downs |
| 1972 | 2 563 | 29 | 818 | 13 | 0.62 | 0.87 | 0.52 | 0.57 |
| 1973 | 1 890 | 17 | 1445 | 11 | 0.50 | 0.71 | 0.50 | 0.53 |
| 1974 | 1 151 | 183 | 902 | 11 | 0.46 | 0.71 | 0.52 | 0.51 |
| 1975 | 699 | 160 | 501 | 8 | 0.52 | 0.81 | 0.50 | 0.53 |
| 1976 | 717 | 24 | 434 | 11 | 0.51 | 0.99 | 0.49 | 0.58 |
| 1977 | 985 | 109 | 507 | 15 | 0.47 | 0.63 | 0.45 | 0.62 |
| 1978 | 1 323 | 216 | 582 | 41 | 0.46 | 0.86 | 0.44 | 0.48 |
| 1979 | 2 370 | 83 | 662 | 162 | 0.48 | 0.63 | 0.44 | 0.46 |
| 1980 | 2 671 | 18 | 540 | 324 | 0.46 | 0.70 | 0.46 | 0.43 |
| 1981 | 2 683 | 27 | 764 | 563 | 0.46 | 0.72 | 0.47 | 0.47 |
| 1982 | 3 132 | 466 | 913 | 592 | 0.46 | 0.63 | 0.45 | 0.41 |
| 1983 | 3 537 | 2 567 | 1 460 | 489 | 0.46 | 0.65 | 0.47 | 0.39 |
| 1984 | 4 420 | 4 038 | 3 496 | 579 | 0.46 | 0.62 | 0.46 | 0.39 |
| 1985 | 6 029 | 4 153 | 4 583 | 747 | 0.46 | 0.61 | 0.45 | 0.39 |
| 1986 | 6 543 | 2 783 | 4 401 | 683 | 0.46 | 0.62 | 0.44 | 0.40 |
| 1987 | 8 079 | 3 500 | 5 817 | 1 047 | 0.46 | 0.62 | 0.45 | 0.39 |
| 1988 | 10 349 | 8 753 | 6 834 | 1 427 | 0.50 | 0.64 | 0.46 | 0.39 |
| 1989 | 10 086 | 5 323 | 4 413 | 2 608 | 0.53 | 0.61 | 0.44 | 0.41 |
| 1990 | 7 399 | 6 126 | 4 798 | 3 061 | 0.55 | 0.62 | 0.50 | 0.45 |
| 1991 | 3 801 | 5 077 | 2 956 | 2 918 | 0.47 | 0.80 | 0.49 | 0.45 |
| 1992 | 2 153 | 2 443 | 811 | 2 028 | 0.51 | 0.81 | 0.54 | 0.44 |
| 1993 | 1 281 | 923 | 765 | 1 938 | 0.68 | 0.88 | 0.55 | 0.43 |
| 1994 | 1 363 | 752 | 705 | 1 409 | 0.63 | 1.32 | 0.57 | 0.43 |
| 1995 | 3 108 | 612 | 585 | 1 058 | 0.54 | 1.32 | 0.65 | 0.44 |
| 1996 | 4 049 | 499 | 678 | 1 763 | 0.56 | 0.86 | 0.66 | 0.39 |
| 1997 | 6 402 | 272 | 635 | 2 802 | 0.58 | 0.89 | 0.72 | 0.41 |
| 1998 | 9 133 | 1 290 | 595 | 2 494 | 0.61 | 0.91 | 0.62 | 0.39 |
| 1999 | 10 182 | 561 | 823 | 2 586 | 0.59 | 0.79 | 0.60 | 0.39 |
| 2000 | 11 305 | 199 | 1 508 | 2 881 | 0.59 | 0.71 | 0.63 | 0.39 |
| 2001 | 13 608 | 637 | 3 189 | 4 233 | 0.61 | 0.79 | 0.64 | 0.39 |
| 2002 | 13 035 | 2 551 | 6 326 | 4 515 | 0.59 | 0.82 | 0.66 | 0.40 |
| 2003 | 11 585 | 5 763 | 9 794 | 8 157 | 0.57 | 0.84 | 0.63 | 0.40 |
| 2004 | 11 392 | 7 816 | 9 705 | 10 067 | 0.57 | 0.85 | 0.66 | 0.40 |
| 2005 | 9 843 | 4 223 | 8 684 | 9 523 | 0.55 | 0.81 | 0.67 | 0.39 |
| 2006 | 8 841 | 1 925 | 6 763 | 11 837 | 0.51 | 0.84 | 0.69 | 0.39 |
| 2007 | 8 517 | 2 974 | 5 579 | 12 851 | 0.56 | 0.82 | 0.75 | 0.40 |
| 2008 | 9 872 | 2 274 | 8 462 | 15 119 | 0.59 | 0.92 | 0.85 | 0.46 |
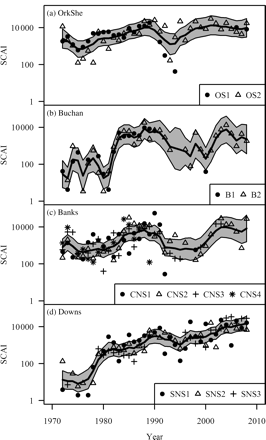
Time-series of modelled SCAIs on a log scale (heavy line), with 95% confidence intervals (grey region) for the (a) Orkney–Shetland (OrkShe), (b) Buchan, (c) Banks, and (d) Downs spawning components, compared with LAI observations (placed on a common ground by dividing by the fitted spawning proportion, pu, for that unit). The key at the bottom-right of each panel relates the symbols to the sampling-unit codes given in Table 1.
Extensions to the model were examined using the likelihood-ratio test. Equation (5) assumes that the precision of the observations is constant across the sampling units within a spawning component. The sensitivity of the model to this assumption was tested by modifying the model to have a unique σLAI for each sampling unit, which meant adding 1–3 additional parameters per component. When considered across all four spawning components, the improvement in the fit was not significant (likelihood ratio = 12.0, d.f. = 7, p = 0.10); therefore, this more complex model was rejected. Similarly, reductions in funding of the survey (and consequently in sampling effort) around 1990 (ICES, 2009) might have reduced the precision of the LAI estimates. Therefore, a version of the model was considered fitting two σLAI parameters, one for observations up to 1989 and one for observations from 1990 onwards. Across all spawning components, there was no significant improvement in model fit (likelihood ratio = 7.9, d.f. = 4, p = 0.10); therefore, this modification was also rejected.
As a test of the model's ability to deal with gaps in the observations, a resampling procedure was used. A small fraction (10%) of the observations were removed randomly (the “test set”) and the remaining data (the “tuning set”) were used to fit the model. The fitted model parameters were then used to predict the values of the test set, and the mean residual (bias) from the actual observations calculated. The procedure was repeated multiple (100) times to characterize the distribution of biases. The model proved to be capable of predicting the missing observations in an unbiased manner (mean log-residuals of prediction: Orkney–Shetland 0.2 ± 0.9, Buchan 0.2 ± 1.2, Banks 0.0 ± 0.9, Downs 0.0 ± 0.8) with a precision comparable with that obtained using the full dataset.
The sum of the SCAIs for the different components can be interpreted as an index of the total abundance of the spawning stock. After log-transformation of both time-series to account for the assumed lognormal error structure, the summed SCAIs were highly correlated with the SSB estimated by the analytical assessment (r2 = 0.90, p < 0.001). The correlation with the second and completely independent SSB series was slightly lower, but still highly significant (r2 = 0.87, p < 0.001; Figure 3). Moreover, in neither case were the slopes significantly different from 1.0 (0.95 ± 0.11 and 0.96 ± 0.12, respectively), suggesting a direct proportionality between the summed SCAI and SSB.
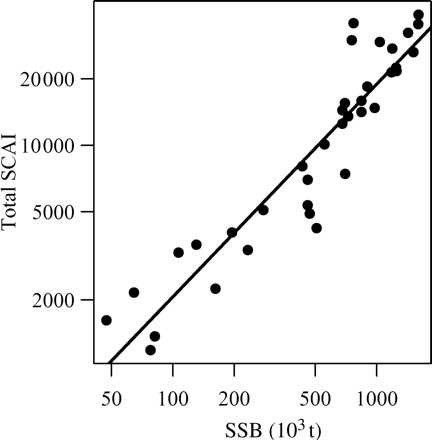
Relationship between the sum of the SCAIs and the SSB from 1972 to 2008 on log scales. The SSB is estimated using the standard assessment of this stock, but excluding the MLAI. The solid line represents a linear regression between the log-transformed values (r2 = 0.87, p < 0.001, slope = 0.96 ± 0.12).
Examination of the pu parameters [Equation (5)] reveals that the spawning intensity varies across sampling units (Figure 4). In several cases, the differences between sampling units within components are significant. Specifically, the Buchan (CNS) and Downs (SNS) components exhibit clear peaks, with spawning intensity trailing off markedly later in the season.
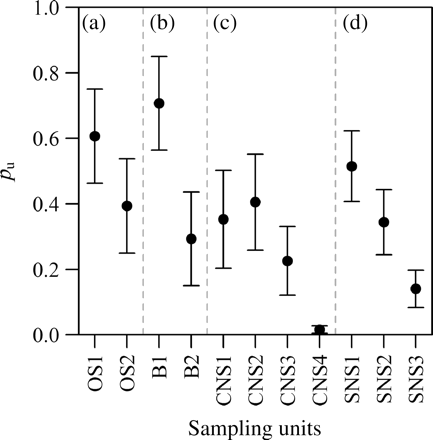
Proportion of the total component spawning intensity, pu, by sampling unit (cf. Table 1) as estimated by the model for the (a) Orkney–Shetland, (b) Buchan, (c) Banks, and (d) Downs spawning components. Error bars represent the 95% confidence interval on the estimated proportion.
There is a broad agreement in the development of the SCAI of the different spawning components over time (Figure 5), although these estimates are completely independent. Each component appears to have generally followed the overall dynamics of the stock, starting from low levels in the 1970s followed by recovery and peaks in the late 1980s and early 2000s. However, there are also appreciable differences. Immediately obvious is that the Orkney–Shetland spawning component appears to have increased much quicker after the two recent depletion events (after 1976 and 1993) than the others did. In contrast, the Downs component remained in a relatively depressed state during the 1980s recovery and has only been rapidly rebuilding during the 2000s. The dynamics of the Buchan and Banks components are quite similar, and generally lie between the extremes represented by Orkney–Shetland and Downs.
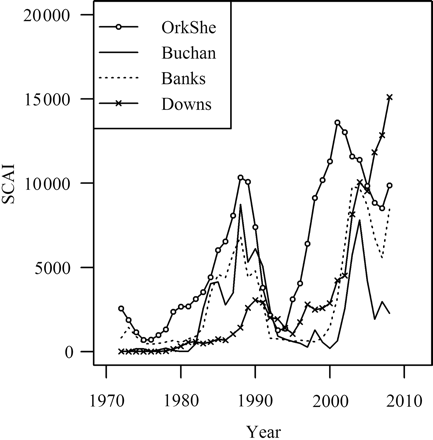
The different dynamics of the components cause large changes in the stock composition (Figure 6). In particular, the faster increase in the Orkney–Shetland component from lows in 1976 and again in 1993 means that the biomass recovery of the overall North Sea herring stock was led by this component. The largest fraction of the total SSB in the past 35 years has generally been represented by the Orkney–Shetland component (on average 50%), but the ratio has ranged between 25 and 80%. In recent years, the Downs component has made a comeback and it now represents the largest component, whereas the Buchan component is now the smallest.
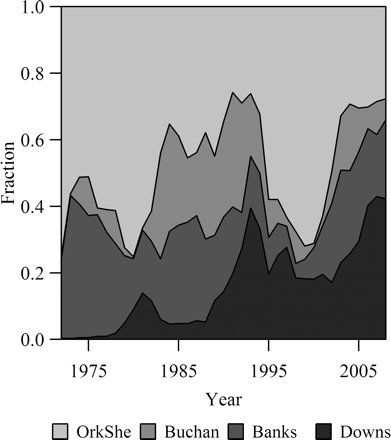
Time-series of the fractional contribution of each spawning component (cf. Figure 2) to the total stock, as estimated from the SCAIs, with shaded areas arranged from top to bottom according to the north-to-south arrangement of the components.
Discussion
This simple state-space model allows for the estimation of trends in the four different components of the North Sea autumn-spawning herring stock. Assessing the quality and appropriateness of such a model, however, remains a challenging task. Here, a number of different approaches have been used, including visual examination of the fit, statistical tests to ensure that the underlying assumptions are met, examination of alternative model configurations, and a comparison with other indices of stock abundance. Perhaps, the most powerful test is the close agreement between the combined indices and completely independent SSB estimates for the entire stock: as each index is fitted independently to data for the single spawning components, this result strongly suggests that the model is capturing the underlying component dynamics as well.
However, several weaknesses can readily be identified in the model. The first is that the assumption of population integrity (assumptions I and II) might not be fulfilled. Herring stocks do sometimes change their spawning grounds (Glover, 1957; Geffen, 2009), and some might even utilize multiple spawning grounds during a lifetime (Slotte, 1999). A change in spawning distribution is known to have occurred in the North Sea, when Aberdeen Bank (a geographical region within the Buchan spawning component; Figure 1) was recolonized in 1983 (Corten, 1999). Nevertheless, the model fits the observations during 1980–1985 well and, therefore, appears to have handled this event gracefully (Figure 2b), suggesting that it is relatively robust to changes of this nature.
Changes in the spawning phenology might also violate the underlying assumptions of the model (specifically assumption III). Natural variability in the timing of spawning around the mean distribution could be expected (e.g. owing to variability in environmental cues), but systematic changes in the phenology are also known in fish stocks (e.g. Baltic cod; Wieland et al., 2000). Natural variability in timing will add noise to the pu parameter and, therefore, be absorbed into the error term in Equation (5): 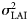 could therefore be thought of more accurately as the sum of noise owing to observation error and to variations in the timing of spawning. Systematic changes in the spawning phenology, however, will have the effect of introducing a time-dependent bias. Such a bias would become apparent in both the pu parameter (in the form of unusually broad confidence intervals) and also as a systematic pattern in the residuals. There is no strong evidence in the results obtained to suggest that such systematic changes have happened in this stock during the period investigated. However, given the relatively low temporal resolution in the survey design and the number of gaps in the data, it is impossible to rule out the possibility completely.
could therefore be thought of more accurately as the sum of noise owing to observation error and to variations in the timing of spawning. Systematic changes in the spawning phenology, however, will have the effect of introducing a time-dependent bias. Such a bias would become apparent in both the pu parameter (in the form of unusually broad confidence intervals) and also as a systematic pattern in the residuals. There is no strong evidence in the results obtained to suggest that such systematic changes have happened in this stock during the period investigated. However, given the relatively low temporal resolution in the survey design and the number of gaps in the data, it is impossible to rule out the possibility completely.
The model uses the readily available LAIs as the source data. However, in using these indices, other information gathered during the IHLS, such as the larval length distributions and catches of larvae >10 mm (>11 mm for the Downs) remain unused. Once available, the use of larval production estimates that incorporate this kind of information might improve the quality of the input further.
The summed SCAIs are closely correlated with the MLAI (r2 = 0.95, p < 0.001), as might be expected, because the two indices simply represent different ways of working up the same time-series of larval abundance. However, the SCAIs have the important advantage that, rather than assuming that the relative distribution among the spawning components is constant over time, each component is treated individually and there is no assumption of correlation between components. In fact, the individual components clearly display appreciable differences in their dynamics and, therefore, in their relative stock composition (Figure 6), invalidating the basic assumption underlying the MLAI. Therefore, although the MLAI appears to work well as an index of overall stock abundance (Simmonds, 2009), it is not a suitable basis for drawing conclusions about the dynamics of individual components.
The SCAI model can cope with the presence of occasional gaps in the time-series of observations. Multiple missing observations (cf. Figure 2b: 1994–1995) are treated in a manner consistent with human intuition, i.e. by interpolating between the neighbouring observations and increasing the uncertainty of the indices at these points. Tests based on a subset of the data demonstrated that the model could both fit the reduced dataset and still make unbiased predictions of the omitted observations. Moreover, the ability to handle missing observations allows a time-series of observations, such as from CNS4 (which was last sampled in 1989 and has since been discontinued) to be used still. Although many modelling approaches would have to discard such time-series, because of their limited extent, this model incorporates all information for all time-series. This robustness to missing observations is a clear advantage for the analysis of patchy data, because the application to IHLS data gives insight into the historic dynamics of the components not available previously.
Regarding design and execution of the IHLS, there appears to be a mismatch between the sampling units most frequently surveyed and those that cover the greater part of the spawning activity. This is especially apparent in the northern components, where the OS1 and B1 units capture 60 and 70% of the spawning, respectively, whereas OS2 and B2 are carried out more frequently (Table 1). The majority of the Downs spawning occurs during the first sampling unit (SNS1), suggesting that it might be worth adding an earlier survey unit (1–15 December) to ensure that the entire spawning period is adequately covered. Finally, the CNS4 unit only covers around 1.5% of the spawning and its abandonment in the early 1990s (ICES, 2006) therefore appears justified.
Although there are common trends among the different spawning components, there are also substantial differences in their dynamics. As a direct consequence, the relative composition of the entire stock changes throughout time, possibly giving rise to differences in exploitation rates (Bierman et al., 2010) and the associated risk of local depletions (Kell et al., 2009). Clearly, maintaining such spatial diversity within a stock should provide resilience to both anthropogenic and natural stressors (Harden Jones, 1968; McPherson et al., 2001; Secor et al., 2009). Continued monitoring of the spatial composition of the North Sea herring stock, through the IHLS survey in conjunction with the type of model advocated here, is critical if management advice is to incorporate measures that could help to avoid local depletions.
The results support and strengthen those of Schmidt et al. (2009), who suggested that a recovery seen in the total SSB of a stock does not necessarily mean that the stock has fully recovered in all its aspects. Figure 6 clearly shows that whereas the total SSB recovered relatively quickly from the 1970s collapse, it was not until at least the early 1990s, and possibly the mid-2000s, before the stock had regained its component diversity.
The approach presented here addresses one side (larval production) of the component dynamics of North Sea herring. The other side is to understand recruitment at the component level. Recent work has established that year-class strength, at the total stock level, is determined between the early larval stage and the late larval stage (Nash and Dickey-Collas, 2005; Payne et al., 2009). Resolving the problem of assigning observations on the distribution of late larvae to the spawning component from which they originate (e.g. using hydrographic-drift models; Christensen et al., 2007) therefore potentially yields not only component-level recruitment information, but also it might help to pinpoint the events contributing to differential survival. In this sense, the SCAI model represents a first step in the direction of a full component-resolved understanding of the dynamics of the North Sea autumn-spawning herring stock.
Acknowledgements
I wish to recognize the crews, captains, and scientists who have taken part in the International Herring Larval Survey over the past 40 years and have been instrumental in collecting this wonderful time-series. I also thank Mark Dickey-Collas, Richard Nash, Uffe Høgsbro Thygesen, Lotte Worsøe-Clausen, and Anders Nielsen for inspiring conversations, encouragement, and constructive criticism. This study was funded by the EU FP6 project UNCOVER (contract number 022717) and the project Defineit under the MariFish ERA-NET (ERAC-CT-2006-025989). A copy of the data, analysis scripts, and SCAI time-series is available from the author or the ICES HAWG stock assessment repository, http://hawg.googlecode.com/.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}