





Abstract
Early hatching has been shown to be associated with increased survival of Norwegian spring-spawning herring (Clupea harengus) larvae. We investigated whether the process behind this association is related to larval drift and ambient temperature. A three-dimensional hydrodynamic model was used to simulate the effect of hatching date on northward displacement and temperature conditions of larvae from spawning grounds off western Norway for 1989–2008. The simulations revealed that the displacement during a period of 60 d was greatest if the larvae hatched early in the season and if they were located near the surface. The relationship between drift speed and ambient temperature was significantly negative, but less so later in the season, because the coastal current became progressively warmer. Results from the simulated interannual variations in larval drift compared with the observed survival suggest that a rapid northward displacement to the main nursery area in the Barents Sea is more important for larval survival than ambient temperature. The significant effect of northward displacement on survival could be explained by reduced overlap with predators and/or higher prey densities, but the causal processes involved remain to be investigated.Vikebø, F. B., Husebø, Å., Slotte, A., Stenevik, E. K., and Lien, V. S. 2010. Effect of hatching date, vertical distribution, and interannual variation in physical forcing on northward displacement and temperature conditions of Norwegian spring-spawning herring larvae. – ICES Journal of Marine Science, 67: 1948–1956.
Introduction
Spawning grounds of Norwegian spring-spawning (NSS) herring (Clupea harengus) are found along the central Norwegian continental shelf (58–69°N), but the main spawning area is located off Møre (62–63°N; Slotte, 2001). Spawning time is variable and ranges from February to April, and the eggs are attached to hard-bottom substrata (gravel, stones, and rocks) down to 250-m depth. The larvae hatch after ∼2–3 weeks, ascend in the water column, and drift with the water masses above 50 m.
Mortality is high throughout the larval stage and subsequently drops substantially (Sætre et al., 2002). Hence, to understand the mechanisms responsible for variations in recruitment, the processes operating during the larval phase are of crucial importance. Husebø et al. (2009) observed a negative correlation between hatching time and recruitment. They suggested that this could be the result of reduced overlap between larvae and their predators in years of early hatching, because predators generally arrive, or become more active, later in spring. If predator activity is linked to temperature, early hatching and a rapid northward drift could be favourable because of the temperature gradient along the coast. However, Husebø et al. (2009) acknowledged that early spawning might have other advantages related to feeding success and growth. Temperature represents one of the important environmental factors affecting growth and survival of fish larvae (Houde, 1989; Morse, 1989). Therefore, it might be clarifying to quantify the direct effect of hatching time on larval drift and on the ambient temperature experienced along the drift route, both within and between years.
The dispersal of early life stages of fish is commonly studied with numerical models because direct observational methods would be a Sisyphean challenge and the costs involved inhibitive. Coupled biophysical models can integrate the states (size, location, survival probability) of individual organisms forward in time, dependent on temporally and spatially varying biotic and abiotic parameters (ICES, 2009a). Such a model approach has previously been used for Northeast Arctic cod (Vikebø et al., 2005, 2007; Opdal et al., 2008), and the same model is used here for NSS herring.
Fiksen et al. (2007) and Vikebø et al. (2007) demonstrated how vertical habitat selection influences drift routes, growth, and survival probability of cod offspring. Observations on cod larvae and juveniles in the area indicated active vertical migration (Ellertsen et al., 1984; Lough and Potter, 1993; Johansen et al., 2009). Although observations on the vertical migration patterns in herring larvae are contradictory (Dragesund, 1970; Bjørke et al., 1986), it seems reasonable to expect that larval feeding and predator avoidance change over time, because both processes depend on light conditions and daylength becomes progressively longer through spring. Hence, the preferred depth-at-age is likely to increase with improving light conditions and hence hatching time.
Our main objectives were to use state-of-the-art models (i) to simulate the effects of hatching date and drift depth on the northward displacement of, and the ambient temperature experienced by, larval NSS herring and (ii) to relate simulated interannual variations in drift and ambient temperature to observations on larval survival (Husebø et al., 2009).
Material and methods
Model description
To assess the importance of temporal and spatial variations in circulation on northward displacement and temperature exposure of larval herring, we utilized a three-dimensional hydrodynamic model (ROMS; Haidvogel et al., 2008; www.myroms.org). Monthly mean lateral boundary conditions were taken from a global version of ROMS, with a resolution of ∼20 km in the North Atlantic and Arctic, and it comprised three-dimensional velocities, sea-surface height (SSH), temperature, and salinity. In addition, eight tidal constituents from the Oregon State University TOPEX/Poseidon Global Inverse Solution (TPXO 7.0; Egbert et al., 1994; Egbert and Erofeeva, 2002) were added to the SSH and barotropic flow at the lateral boundaries. Six-hourly vertical boundary conditions were taken from ERA40 interim (www.ecmwf.int), namely sea-surface air pressure, windstress, latent, sensible, downward shortwave radiative, and net longwave radiative heat flux. The model domain covered the Nordic seas (North Sea, Barents Sea, Kara Sea, and partly the Arctic; Figure 1) and had a spatial resolution of 4 km, with 30 s coordinates (stretched, terrain following) in the vertical. Daily averages were stored on disk for utilization in the offline particle-tracking model LADIM (Lagrangian advection and diffusion model; Ådlandsvik and Sundby, 1994) transporting annually 30 000 particles from the main spawning ground at Møre (Figure 1, black box in bottom right panel) from 1989 to 2008.
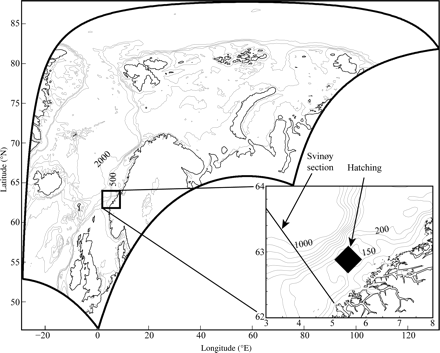
Model domain covering the Nordic seas, North Sea, Barents Sea, Kara Sea, and partly the Arctic Ocean. Contours indicate depths. Inset shows the hatching location at Møre (black diamond) used in the simulations, with the Svinøy cross section (straight black line).
Consistent with recommendations by ICES (2009a), the particle-tracking algorithm utilizing daily mean velocity and temperature fields from the general circulation model (GCM) is state-of-the-art, including a fourth-order Runge–Kutta advection scheme and bilinear three-dimensional interpolation of daily mean velocity and temperature fields. Tides are part of the ocean simulation, but these are averaged out when stored on disk. This is likely to affect dispersal features in the simulations; so, artificial isotropic diffusion in the form of so-called random walk (randomly selected numbers) is added to the advection terms to compensate for the lack of both tidal and subgrid scale mixing. It is scaled in accordance with the horizontal resolution (Ådlandsvik and Sundby, 1994) and set to 20 m2 s−1.
Two different simulation setups were designed for the particle tracking to meet our objectives. First (Experiment A), to study the combined effects of hatching date and drift depth on northward displacement and temperature exposure, we initiated 30 000 particles evenly distributed across the depths 1, 5, 10, 20, 30, and 40 m for all days starting 5 March and ending 25 April (observed early and late thresholds for hatching; Fossum, 1996). Note that all particles were kept at fixed depths and allowed to drift for 60 d before processing the individual displacements and temperature exposures. Second (Experiment B), to simulate the actual drift routes and temperature exposure of larvae in relation to hatching date, all 30 000 particles were initiated at 10 m and allowed to perform an imposed diel migration. An imposed diel migration was also used in simulations of the interannual variations in mean northward displacement and temperature exposure. To ensure discrete periods, early-, mid-, and late-hatched larvae were defined as being hatched between 15 and 20 March, 28 March and 2 April, and 10 and 15 April, respectively, with early and late periods well inside the observed historical range.
The numerical diel vertical migration imposed represents an ad hoc formulation based on observations. Dragesund (1970) established that larval herring were located between 5 and 20 m during night-time and 20 and 40 m during daytime. In our formulation, the larvae actively sought these two depth ranges, depending on light availability and individual swimming capacity set at one-tenth of the body length per second (Vikebø et al., 2007; Kristiansen et al., 2009). A larval herring is assumed to grow 0.5 mm d−1 (Stenevik et al., 1996) from a hatching length of 5 mm. Therefore, a newly hatched larva can migrate vertically at a rate of 1.8 m h−1, whereas a 60-day-old larva (35 mm long according to the simplified growth function above) can migrate at 12.6 m h−1.
Oceanographic features and model validation
Circulation features along the larval drift routes from the spawning grounds to the nursery grounds in the Barents Sea are mainly determined by the relatively warm Norwegian Atlantic Slope Current (NASC) along the shelf break (Orvik et al., 2001; Orvik and Skagseth, 2005) and the cool, less saline Norwegian Coastal Current (NCC) between the NASC and the coast (Sætre, 1999, 2007), both flowing northwards. Current measurements in the Svinøy section outside Møre (Orvik et al., 2001) indicate a stronger NASC during winter (7 Sv; 1 Sv is 106 m3 s−1) than during summer (3 Sv). According to Blindheim et al. (2000), the NASC is locked to the shelf break, but can be displaced off the shelf and weakened with a low North Atlantic Oscillation (NAO) index (Hurrell et al., 2003), or onto the shelf and enhanced with a high NAO. Current velocities are normally between 0.2 and 0.5 m s−1, depending on the time and location, but velocities >1 m s−1 have been observed (Orvik et al., 2001). Based on satellite-tracked drifter buoys (Sætre, 1999), the highest velocities in the NCC were found in the main inner drift route along the deep end of the 200-m isobath close to the coast, followed by an outer drift route at the shelf break, with slightly lower velocities. In between these two routes, the drifters exhibited enhanced residence time, caused by eddies on the top of topographical features, such as banks and troughs. The vertical structure of the NCC transforms from a well-mixed water mass with a strongly tilted interface towards the NASC during winter to a stratified water mass with a weakly tilted front during summer (Sundby, 1984; Sætre, 2007). The seasonal changes in the windfield (strong and mostly southwesterly during winter; less strong and from variable directions during summer) imply a transition from a strong baroclinic flow during winter to a weakened and less unidirectional flow towards summer.
The simulations reproduced hydrographic and circulation features in the Svinøy section (0°E 65°N to 5°E 62.3°N) as described above (Figure 2). The NASC is seen as a strong, north-flowing current over the shelf break, with subsurface temperature and salinity maxima. The slopes of the isohalines at the interface between NCC and NASC are tilted more in March than in June, indicating a seasonal weakening in the NCC. At the same time, the NCC becomes warmer, increasing from <6°C in March to >10°C in June.
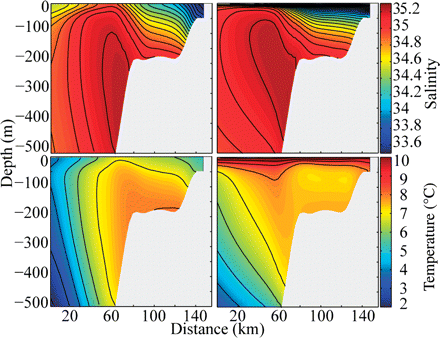
Simulated data from the Svinøy cross section used for model validation: average salinity in March (upper left) and June (upper right) and average temperature in March (lower left) and June (lower right), 1989–2008.
Modelled annual mean volume flux of NASC (defined by temperatures above 5°C and salinity above 35 psu) for 1997–1999 was 4.2 Sv, identical with the values obtained by Orvik et al. (2001) for the same period. There was a strong seasonal signal in the volume transport, with a maximum in winter and a minimum in early summer (Figure 3), consistent with direct observations. In addition, the winter maximum and summer minimum were comparable with observations. The second peak in volume flux during autumn, as evident in the measurements, was captured by the model, but somewhat overestimated.
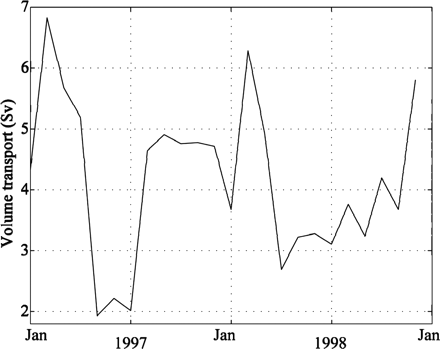
Simulated volume flux (Sv) of the NASC (defined as water of >5°C and >35.0 psu) at the Svinøy cross section.
Figure 4 shows monthly averaged velocities at 20 m, along with temperatures for April 1989–2008 and the corresponding values for 1994, 1995, and 2004. Note that these three years represent outliers with respect to average latitude and temperature conditions after 60 d of drift. Several interesting features appear in these panels. Similar eddy structures in the NASC along the shelf break have been reported by Rossby et al. (2009), using Rafos floats, and by Orvik et al. (2001), using a vessel-mounted Acoustic Doppler Current Profiler. The 1989–2008 average compared with yearly averages indicates that these eddy structures were not persistent and that their locations might vary over time. On the shelf, the core of the NCC was cooler than farther off the coast. The main inner drift route was along the 200-m isoline, as reported in Sætre (1999), with less unidirectional flow above the bank structures, as was also evident from drift experiments. The distance between the core of the NCC and the coastline increased when approaching the Vestfjord, leaving a progressively increasing area of waters with enhanced retention.
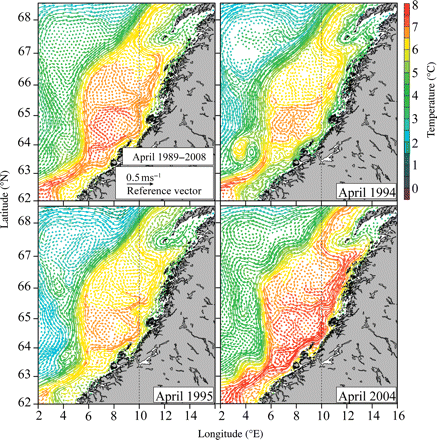
Simulated mean circulation patterns and temperature at 20 m deep during April averaged over 1989–2008 (upper left) and for 1994 (upper right), 1995 (lower left), and 2004 (lower right). The years 1994 and 1995 were characterized by relative low velocities, whereas 2004 was a year with high velocities.
Results
Experiment A
The simulations demonstrated that northward displacement of larvae is influenced by hatching date and drift depth. There was a clear trend towards faster transport of larvae hatched earlier in the season and drifting at shallower depths, resulting in a span of up to 3° latitude in the downstream direction along the Norwegian coast (Figure 5a). In fact, larvae hatched in early March would, on average, be ∼1.5° farther north after 60 d of drift than larvae hatched in late April, when integrated over every fixed drift depth (p < 0.001, r2 = 0.93; Figure 5b). Similarly, drift at different depths of the upper 40 m will on average result in ∼1.5° difference in the latitudinal downstream direction (p < 0.001, r2 = 0.84). The increase in average latitude with decreasing drift depth implies a stronger shear near the surface than farther down in the water column, consistent with flow in the near-surface Ekman layer.
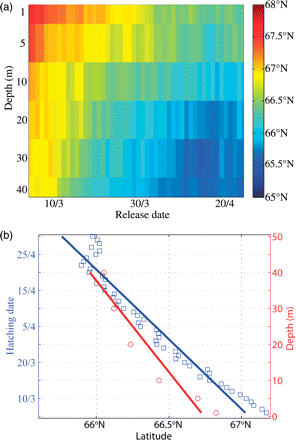
Characteristics of particles after 60 d of drift for 1989–2008: (a) mean latitude as a function of fixed depth and hatching date; and (b) effects on northward displacement when integrating hatching date over every fixed drift depth (blue) and when depth is integrated over every hatching date (red).
Effects of fixed drift depth and hatching date on the northward displacement varied between years (Figure 6). Although most years were consistent with the average pattern (Figure 5a), some years displayed aberrant patterns. In 1996 and 2000, effects of drift depth and hatching date were virtually non-existent, and the northward displacement increased rather than decreased with hatching date in 2002 and 2006.
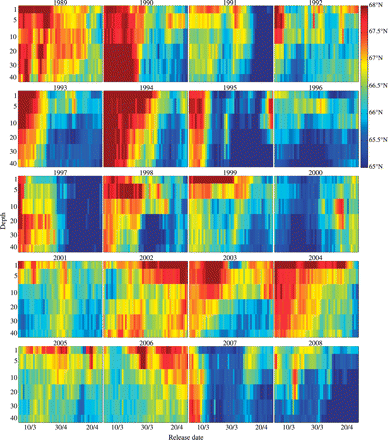
Average latitude reached after 60 d of drift as a function of fixed depth and day of hatching, 1989–2008.
Experiment B
By averaging the temperature exposure during the 2 months of drift for all particles ending up at different latitudes by year for early-, mid-, and late-hatched larvae, respectively, we obtained three curves for each year with temperature as a function of latitudinal endpoint. These sets were then averaged over the different years by hatching date and the standard errors were calculated. An ANOVA demonstrated that the mean temperature experienced over the period of 60 d was significantly influenced by the latitudinal endpoint (p < 0.001) and by the hatching date (p < 0.001), decreasing northwards from 62°N and increasing with delayed hatching. However, the negative temperature effect of northward displacement tended to fade north of 67°N (Figure 7). When running an ANOVA using only particles ending up in the area 67–70°N, the latitudinal endpoint did not have a significant effect on the temperature experienced. In addition, the shape of the year-specific curves (not illustrated) varied substantially. Years of slow drift (1995 and 1997; cf. Figure 4) displayed less temperature difference between particles ending up at different latitudes, whereas years with strong drift displayed the largest difference.
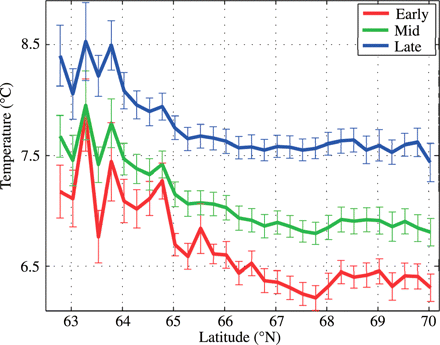
Mean ambient temperature (means of year-specific means with standard errors) encountered after 60 d of drift as a function of latitudinal endpoint for early-, mid-, and late-hatched larvae.
The mean simulated drift trajectories of larvae (averaged over all years) ending up south and north of 68°N after 60 d of drift, as well as the temperatures experienced at different latitudes, were similar for the early-hatched larvae. However, the mid- and late-hatched larvae ending up north drifted farther from the coast and they experienced lower temperatures (Figure 8). Therefore, ANOVAs proved that the ambient temperature along the drift route for mid- and late-hatched larvae increased with latitude up to 68°N (p < 0.001), and the ambient temperature was lower for larvae ending up north than those ending up south (p < 0.001). Conversely, the ambient temperature for early-hatched larvae decreased significantly (p < 0.001) for both groups (no significant difference). The simulations indicated that larvae undergoing a rapid northward transport are either early-hatched larvae that drift in the core of the NCC or mid- and late-hatched larvae that drift on average farther offshore, whereas the slow-moving larvae are confined to the core of the NCC. However, output that is more detailed indicated that the fast-drifting larvae could use either a drift route in the NCC or one along the shelf break, with not much drifting in between. Therefore, the mean trajectory of the fast-moving larvae probably has little biological relevance, but reflects the fraction drifting along the shelf break. The increasing distance with hatching date between the two mean trajectories therefore probably indicates that relatively more fast-moving larvae are drifting along the shelf break. On average, the slow-moving larvae spent ∼20–30 d longer to reach 67°N than the faster-drifting larvae.
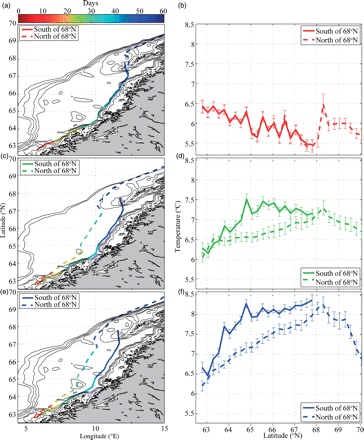
Average drift route and northward displacement of larvae as a function of day (indicated by colour), for individuals ending up north (dashed) or south (solid) of 68°N, and for (a) early-, (c) mid-, (e) and late-hatched individuals and the corresponding mean ambient temperatures experienced (with standard errors based on variations in year-specific means) at different latitudes along the drift route (b, d, and f).
The mean latitude reached after 60 d of drift and the integrated mean temperature exposure during this period varied substantially among years for early-, mid-, and late-hatched larvae (Figure 9a and b). The mean latitude reached ranged from 63.8 to 68.1°N, with early-hatched larvae in general ending up farther north than those hatched later in the season. The mean temperature exposure ranged between 5.3 and 8.0°C, and generally increased with hatching date. According to the simulations, larvae were exposed to very low temperatures in 1994, co-occurring with a high latitudinal distribution after 60 d. Larvae in 2004 were also distributed far north after 60 d of drift independent of hatching date, but they experienced relatively high temperatures (cf. Figure 4). These findings are summarized in Figure 9c by plotting the average value for early-, mid-, and late-hatched larvae, along with blue circles indicating years of above-median survival (recruitment at age 0 divided by spawning-stock biomass), based on ICES (2009b). All 9 years of enhanced survival are located in the top right quadrant, characterized by mean ambient temperatures >6.7°C and latitudes higher than 66°N, suggesting that both features are important in respect of potential survival, although there are also three low-survival years in this quadrant. Most years of poor survival (seven out of ten) were located outside this quadrant. Six had apparently favourable ambient temperature conditions without enhanced survival, whereas only 4 years had favourable latitudinal distribution without enhanced survival. This suggests that latitude is more important for survival than temperature. Although the estimates of survival during years after 2004 are uncertain because data from the fishery are lacking, all survey information indicates that these were small year classes and that survival has been low since 2004.
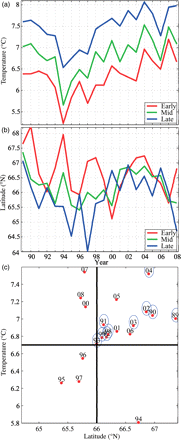
Average (a) temperature and (b) latitude by year after 60 d of drift for early-, mid-, and late-hatched larvae, along with (c) a scatterplot of average temperature vs. latitude (for early-, mid-, and late-hatched larvae combined), 1989–2008. Blue circles indicate enhanced survival (above median; adapted from ICES, 2009b). The cross is defined by 6.7°C and 66°N.
Discussion
The results suggest that the GCM can capture the oceanographic features that are important for the drift and temperature exposure of the early-life stages of NSS herring. An experiment has been designed, and will be carried out soon, to validate further the model by determining its predictive skill by comparing modelled drift and recaptures of drifters at sea. We expect the results to contribute to the further development of tools to study drift trajectories in the early-life history of pelagic species.
The simulations in Experiment A demonstrated that northward displacement of larvae was generally faster if hatching was early in the season, as a consequence of the winds becoming weaker and less unidirectional through spring, resulting in weaker northward flows along the coast. In addition, drift at a shallow depth resulted in a faster northward transport than drift at greater depths. In Experiment B, the strong negative relationship between northward drift speed and ambient temperature became weaker later in the season as the coastal current became progressively warmer. However, results from the simulated interannual variations in larval drift compared with observed survival suggest that a rapid northward displacement in relatively warm water, to the main nursery area in the Barents Sea, is favourable for survival, although quick transport seems more important for larval survival than the ambient temperature. Hence, years of anomalously strong northward transport seem to add to the positive effect of early hatching on survival suggested by Husebø et al. (2009).
In reviewing the literature, Wright and Trippel (2009) found several examples of fish species exhibiting selection for either early or late hatching and they concluded that the timing of reproduction on its own could be important for offspring survival. Husebø et al. (2009) ascribed the observed negative relationship between survival and hatching date in NSS herring to less overlap with predators early in the season, and consequently less predation. This has also been demonstrated for several other species, although the early larvae experience lower temperatures and exhibit lower growth rates (Fortier and Quinonez-Velazquez, 1998; Lapolla and Buckley, 2005; Nishimura et al., 2007).
Husebø et al. (2009) observed that the relationship was less pronounced from 2000 on and that this change was related to a northward extension of the spawning grounds (Slotte, 2001). In our experiments, all particles were released on the southern spawning grounds off Møre, but part of the stock is now spawning north of 63°N (up to 70°N). The larvae originating in the northern spawning grounds have a much shorter drift route to the nursery areas in the Barents Sea. Therefore, the simulations are not representative of the total stock, although the greater part still utilizes the Møre spawning grounds.
The simulations did not include year-specific differences in hatching date, distribution of post-hatch larvae, or predator-and-prey distribution. Previous studies focusing on match–mismatch with both predators and prey (Fossum and Moksness, 1993; Fossum, 1996; Husebø et al., 2009) indicated that this is a difficult theme to cover because of a lack of data. However, extensive data on hatching date and larval distribution are available and these could be included in future applications of the model. Now that the effects of hatching date and drift depth on northward displacement have been demonstrated, it seems appropriate to model the real annual drift based on observed hatching curves and distribution along the coast. In addition, the focus should be on aggregating all available data on prey and predator distributions, and possibly on conducting dedicated surveys to collect relevant data, to understand fully the processes behind the observed relations between hatching date, northward displacement, and survival through early life stages in NSS herring.
Acknowledgements
The study was carried out with financial support from the Commission of the European Communities as a contribution to FP6 Specific Targeted Research Projects 044133 (RECLAIM) and 022717 (UNCOVER). The paper does not necessarily reflect the views of the Commission. The authors are indebted to the guest editor, Niels Daan, for valuable suggestions on how to improve the manuscript.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}