





Abstract
Realized habitats of North Sea herring for two larval and two juvenile stages were estimated over the past 30 years, using abundances from surveys tied to modelled estimates of temperature and salinity. Newly hatched larvae (NHL) were found mainly in water masses of 9–11°C, pre-metamorphosis larvae (PML) around 5–6°C, juveniles aged 0 in summer around 13–14°C, and juveniles aged 1 in winter around 4–5°C. The median salinity in which the NHL were distributed was 34.4–35.0 and 33.7–33.9, respectively, for PML and juveniles. Interannual variations in temperature and geographic variables in the North Sea were compared with the time-series of realized habitats. The realized temperature habitats of the NHL did not change over time, but the habitat of juveniles in summer may be associated with higher temperatures. Juveniles aged 1 in winter are found in waters colder than the average for the North Sea, a result also reflected in their geographic shift east into shallower water. The results suggest that juveniles could be limited by temperature, but may also track changes in food or predator distribution, and/or internal population dynamics. Time-series analysis of realized salinity habitats was not possible with the available data because of differences between model outputs.Röckmann, C., Dickey-Collas, M., Payne, M. R., and van Hal, R. 2011. Realized habitats of early-stage North Sea herring: looking for signals of environmental change. – ICES Journal of Marine Science, 68: .
Introduction
The idea of defining habitats for pelagic fish has become more developed in recent years (van der Lingen et al., 2005), partly as a tool to provide quantitative evaluations in response to rising concerns about the impact of climate change on fish populations (Drinkwater, 2005; Rose, 2005; Rijnsdorp et al., 2009). Here, we expand the concepts of spawning habitats to the early life stages of pelagic fish.
Van der Lingen et al. (2005) described the operational concepts of “potential”, “realized”, and “successful” spawning habitats. That approach is subtly different from the common and legal concept of “essential fish habitat” (ICES, 2008), which is an area critical to the long-term survival and health of fish populations (Valavanis, 2008). Successful spawning habitat is the habitat from which the surviving generation is produced; of the three listed by van der Lingen et al. (2005), this is most like the concept of essential fish habitat. The potential habitat with respect to a given variable is defined by the inherent requirements of the fish, so represents the region of space (and time) where the given variable is within appropriate limits for the life stage of the fish to survive. Here, however, we focus on the realized habitat of early life stages of North Sea autumn spawning herring (Clupea harengus). This is the habitat where the fish are actually observed and can be defined with respect to one or more variables (including space and time dimensions). The interaction of connectivity, ecosystem productivity, exploitation, and population dynamics can influence the distribution of fish and hence the realized habitat (ICES, 2008).
Given the large number of variables in this multidimensional space, we expect that some variables will directly constrain the realized habitat, and some will not. Changes in the environment, both as variability around a mean or as a trend, can potentially alter the realized habitat. For a particular environmental variable that is not constraining, changes in the corresponding realized habitat will follow changes in the environment. Alternatively, if that variable is constraining, the corresponding realized habitat will not change because it is the only region (in variable space) suitable for the fish. Examining the relationship between an environmental variable and the corresponding realized habitat can therefore be used as a way of identifying whether or not a variable is constraining.
North Sea herring are close to the centre of the meridonal distribution of herring in the Northeast Atlantic (Blaxter and Hunter, 1982). It would therefore be assumed that the recent changes in the physical environment of the North Sea (Edwards et al., 2002, 2007; Leterme et al., 2008; Drinkwater, 2010) would have limited effect on their overall distribution. However, the productivity of North Sea herring has changed recently (Nash and Dickey-Collas, 2005; Payne et al., 2009), which may be reflected in changes in their distribution and realized habitat.
The characteristics and the locations of North Sea herring spawning/hatching and nursery grounds are well described (Cushing and Bridger, 1966; Burd and Howlett, 1974; Burd, 1978; Heath et al., 1997; Schmidt et al., 2009; Payne, 2010; Figure 1), but the realized physical environment of larval and juvenile stages is not (Heath et al., 1997). Scientific knowledge of North Sea herring is good in the sense that for more than 30 years, five research surveys have been carried out regularly each year, targeting, through survey selectivity by gear, time, and spatial coverage, different ontogenetic stages of herring (Heessen et al., 1997; Simmonds, 2009). Here, we analyse the time-series of survey indices of two larval and two juvenile herring stages (Table 1). Outputs from biophysical models are used to provide temporal and spatial estimates of the physical environment of the North Sea (Skogen et al., 1995; Skogen and Søiland, 1998; Schrum et al., 2006a, b). By linking these two sources of data, proxies for the realized habitat of herring at specific stages of their life can be determined.
The herring survey time-series, by life stage, used in the current study (see Figure 1 for the spatial distributions of the surveys).
| Life stage | Survey | Period | Spatial sampling |
|---|---|---|---|
| NHL | International herring larvae survey | 1972–2006 | Fixed point on sampling grid |
| PML | IBTS plankton samples, quarter 1 (winter) | 1977–2004 | Fixed point within ICES statistical rectangle |
| Juveniles aged 0 (J0) | IBTS quarter 3 (summer) | 1991–2006 | Random location in each ICES statistical rectangle |
| Juveniles aged 1 (J1) | IBTS quarter 1 (winter) | 1965–2007 | Two random locations in each ICES statistical rectangle |
| Life stage | Survey | Period | Spatial sampling |
|---|---|---|---|
| NHL | International herring larvae survey | 1972–2006 | Fixed point on sampling grid |
| PML | IBTS plankton samples, quarter 1 (winter) | 1977–2004 | Fixed point within ICES statistical rectangle |
| Juveniles aged 0 (J0) | IBTS quarter 3 (summer) | 1991–2006 | Random location in each ICES statistical rectangle |
| Juveniles aged 1 (J1) | IBTS quarter 1 (winter) | 1965–2007 | Two random locations in each ICES statistical rectangle |
IBTS, International Bottom-Trawl Survey; NHL, newly hatched larvae; PML, pre-metamorphosis larvae.
The herring survey time-series, by life stage, used in the current study (see Figure 1 for the spatial distributions of the surveys).
| Life stage | Survey | Period | Spatial sampling |
|---|---|---|---|
| NHL | International herring larvae survey | 1972–2006 | Fixed point on sampling grid |
| PML | IBTS plankton samples, quarter 1 (winter) | 1977–2004 | Fixed point within ICES statistical rectangle |
| Juveniles aged 0 (J0) | IBTS quarter 3 (summer) | 1991–2006 | Random location in each ICES statistical rectangle |
| Juveniles aged 1 (J1) | IBTS quarter 1 (winter) | 1965–2007 | Two random locations in each ICES statistical rectangle |
| Life stage | Survey | Period | Spatial sampling |
|---|---|---|---|
| NHL | International herring larvae survey | 1972–2006 | Fixed point on sampling grid |
| PML | IBTS plankton samples, quarter 1 (winter) | 1977–2004 | Fixed point within ICES statistical rectangle |
| Juveniles aged 0 (J0) | IBTS quarter 3 (summer) | 1991–2006 | Random location in each ICES statistical rectangle |
| Juveniles aged 1 (J1) | IBTS quarter 1 (winter) | 1965–2007 | Two random locations in each ICES statistical rectangle |
IBTS, International Bottom-Trawl Survey; NHL, newly hatched larvae; PML, pre-metamorphosis larvae.
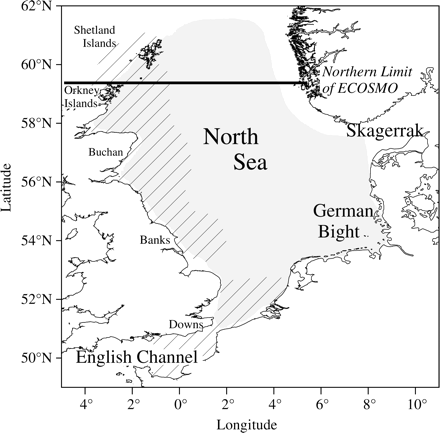
The spatial distribution of survey data. The hatched area shows the location of NHL surveys, and the shaded area locations of PML, J0, and J1 surveys. The northernmost limit of the biophysical model ECOSMO is also shown. Abbreviations are listed in Table 1.
We have two main objectives: (i) to provide estimates of the characteristics of realized habitat of early-stage North Sea herring in terms of temperature, salinity, depth, and geographic coordinates; and (ii) to determine whether these realized habitats change with time and have been or are likely to be influenced by environmental trends, and hence future climate change. Time-series of temperature and salinity are compared with the time-series of realized habitats of larval and juvenile herring.
Material and methods
The realized habitat for herring larvae and juveniles was described quantitatively by combining the abundance estimates of North Sea autumn and winter spawning herring surveys (Table 1) with data on key environmental variables modelled in space and time by two biophysical models (Table 2).
Specifications of the output datasets employed from the biophysical models ECOSMO and NORWECOM.
| ECOSMO | |
| Temporal resolution | Weekly averages |
| Temporal coverage | 1 January 1958 to 31 December 2004 |
| Spatial resolution | ∼10 km × 10 km on the model regular spherical coordinate grid |
| Spatial coverage: North Sea domain | Latitude: 49.3 to 59.0, Longitude: −3.7 to 10.0 (decimalized degrees) |
| NORWECOM | |
| Temporal resolution | Monthly averages |
| Temporal coverage | 1 January 1985 to 31 December 2005 |
| Spatial resolution | 10 km × 10 km on the model polar-stereographic grid, 21 σ-layers |
| Spatial coverage: North Sea domain | Latitude: 49.4 to 62.0, Longitude: −3.9 to 8.5 (decimalized degrees) |
| ECOSMO | |
| Temporal resolution | Weekly averages |
| Temporal coverage | 1 January 1958 to 31 December 2004 |
| Spatial resolution | ∼10 km × 10 km on the model regular spherical coordinate grid |
| Spatial coverage: North Sea domain | Latitude: 49.3 to 59.0, Longitude: −3.7 to 10.0 (decimalized degrees) |
| NORWECOM | |
| Temporal resolution | Monthly averages |
| Temporal coverage | 1 January 1985 to 31 December 2005 |
| Spatial resolution | 10 km × 10 km on the model polar-stereographic grid, 21 σ-layers |
| Spatial coverage: North Sea domain | Latitude: 49.4 to 62.0, Longitude: −3.9 to 8.5 (decimalized degrees) |
Specifications of the output datasets employed from the biophysical models ECOSMO and NORWECOM.
| ECOSMO | |
| Temporal resolution | Weekly averages |
| Temporal coverage | 1 January 1958 to 31 December 2004 |
| Spatial resolution | ∼10 km × 10 km on the model regular spherical coordinate grid |
| Spatial coverage: North Sea domain | Latitude: 49.3 to 59.0, Longitude: −3.7 to 10.0 (decimalized degrees) |
| NORWECOM | |
| Temporal resolution | Monthly averages |
| Temporal coverage | 1 January 1985 to 31 December 2005 |
| Spatial resolution | 10 km × 10 km on the model polar-stereographic grid, 21 σ-layers |
| Spatial coverage: North Sea domain | Latitude: 49.4 to 62.0, Longitude: −3.9 to 8.5 (decimalized degrees) |
| ECOSMO | |
| Temporal resolution | Weekly averages |
| Temporal coverage | 1 January 1958 to 31 December 2004 |
| Spatial resolution | ∼10 km × 10 km on the model regular spherical coordinate grid |
| Spatial coverage: North Sea domain | Latitude: 49.3 to 59.0, Longitude: −3.7 to 10.0 (decimalized degrees) |
| NORWECOM | |
| Temporal resolution | Monthly averages |
| Temporal coverage | 1 January 1985 to 31 December 2005 |
| Spatial resolution | 10 km × 10 km on the model polar-stereographic grid, 21 σ-layers |
| Spatial coverage: North Sea domain | Latitude: 49.4 to 62.0, Longitude: −3.9 to 8.5 (decimalized degrees) |
Empirical data
Four surveys together cover the entire North Sea herring stock, including the winter spawning grounds in the English Channel (Figure 1). Data from the Skagerrak (north of 55.5°N and east of 8°E) were excluded from our analysis to preclude potential interference from the Western Baltic spring-spawning herring stock.
We consider data that describe two life stages of herring larvae and another two of juvenile herring: NHL are specifically targeted by the international herring larvae surveys, and the other stages are sampled during the international bottom-trawl surveys of the North Sea (Table 1).
newly hatched larvae (NHL), defined as larvae 7–15 mm long, stages 1 and 2 (Doyle, 1977);
pre-metamorphosis larvae (PML), defined as larvae 25–40 mm long, stage 4 (Doyle, 1977);
Juveniles aged 0 (J0) in summer;
Juveniles aged 1 (J1) in winter.
Modelled data
Time-series of the environmental variables bottom temperature (T) and salinity (S) were obtained from the biophysical models ECOSMO (Schrum et al., 2006a, 2006b) and NORWECOM (Skogen et al., 1995; Skogen and Søiland, 1998; Table 2). As the North Sea is well-mixed in autumn and winter owing to thermal convection (Janssen et al., 1999), bottom salinity and temperature can be considered good proxies for the distribution of average salinity and temperature.
ECOSMO's northerly extension is 59°N, so it does not cover the spatial boundaries of the Orkney/Shetland spawning ground (Figure 1). Therefore, only NORWECOM data can be used for the Orkney/Shetland spawning component of NHL. Model data (model grid cell ∼10 × 10 km) were coupled to survey data based on the haul position (start). The week/month in which the sample was taken was coupled to the weekly (ECOSMO) or monthly (NORWECOM) offline model outputs.
Environmental data were partitioned into intervals (bins) by rounding to the nearest degree for temperature (°C), latitude, and longitude and to the nearest multiple of 0.5 for salinity. The choice of bin size influenced the results. With lower resolution (i.e. a larger bin size), the data became too aggregated to yield detail in the results. With higher resolution (i.e. a smaller bin size), there were spurious patterns caused by low sample sizes. The bin sizes chosen represented a subjective balance between these two constraints.
Habitat description
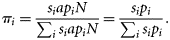
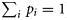 , so given a set of observed fractions for each bin, πi, and the number of samples taken in each bin, si, this series of equations can be solved in a matrix formulation to yield pi as follows:
, so given a set of observed fractions for each bin, πi, and the number of samples taken in each bin, si, this series of equations can be solved in a matrix formulation to yield pi as follows: 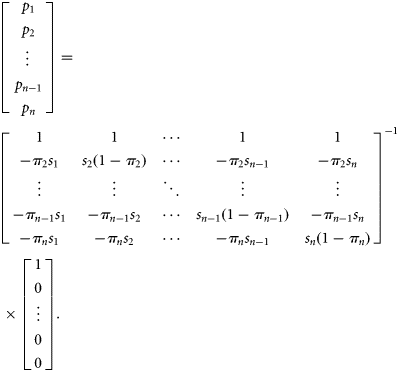
The distribution functions, pi, were summed to yield cumulative distribution functions (CDFs; Perry and Smith, 1994). Annual distribution functions and the subsequent CDFs were calculated by applying the same method for each year separately to investigate potential trends over time (cf. Castonguay et al., 1999). Based on the CDF for a given variable in a given year, we identified the environmental variable value corresponding to the lower quartile (25%), median (50%), and upper quartile (75%) of the distribution of each life stage. We also calculated the average and the standard deviation of all annual medians. If values were missing because the CDF started higher than the median or the quartile, the missing value was replaced with the minimum value.
The variables examined were bottom temperature and bottom salinity. For PML, J0, and J1, which are sampled over the whole North Sea, we also analysed the geographic variables latitude, longitude, and water depth. The spatial analysis was not carried out for NHL because sampling is spatially and temporally limited.
Trend comparison
Subsets of the time-series of environmental data were extracted to compare them with the realized habitats of herring. These were limited in space and time to the coverage of the surveys of NHL, i.e. the spawning grounds, and for the other stages to the whole North Sea at the time of the surveys.
The trend in the annual median distribution values was assumed to represent the trend in the realized habitat. The upper and the lower quartiles of the distribution were used to represent the breadth of the observed habitat. To aid in observing the trends visually, Lowess smoothers (locally weighted scatterplot smoothing; Cleveland, 1979) were applied to the annual medians of the distribution (smoother factor f = 0.5).
To compare whether the annual estimates of realized habitat and the annual environmental signal showed the same trend, a simple linear regression was performed following Douglass et al. (2007). With this approach, a significant slope parameter indicates that two time-series are positively or negatively correlated and that they show a similar or an opposite trend, respectively.
All calculations were programmed in R version 2.7.1. The R scripts and data are available from the authors on request.
Results
Consistency between ECOSMO and NORWECOM
For temperature, the analysis suggests that the two models are consistent (Figure 2a). There is, however, an appreciable discrepancy in the salinity habitats generated from the two (Figure 2b): NORWECOM suggests a wider range in salinity and often yields lower estimates than ECOSMO (Figure 2).
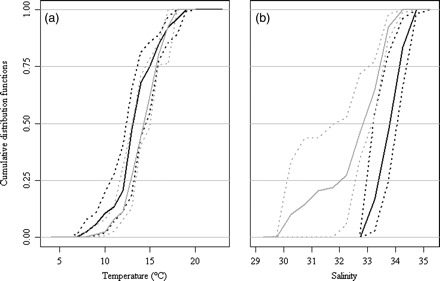
An example comparing the results obtained using the biophysical models ECOSMO (black) and NORWECOM (grey), using the CDFs for life stage J0: median (heavy line), lower and upper quartiles (dotted lines). (a) Temperature and (b) salinity.
Hereafter, temperature and salinity habitats are based mostly on ECOSMO, because ECOSMO has the higher temporal resolution (weekly) and the longer temporal coverage (from 1958 to 2004). NORWECOM data were only used for the Orkney/Shetland spawning component.
Realized habitats of herring
Across all spawning sites, NHL had a realized temperature habitat of 6.1°C (Downs, lower quartile) to 15.1°C (Banks, upper quartile; Table 3). The average of the annual medians of larvae from the three northern spawning grounds was similar (∼11°C, s.d. ∼1.0°C), whereas that from the Downs spawning ground was colder (9.3°C, s.d. 1.2°C; Table 3). The realized salinity habitat of the larvae ranged from 32.6 (Downs, lower quartile) to 35.1 (Orkney, Buchan, and Banks, upper quartile), with an average annual median of 34.9/35.0 for the three northern spawning grounds and 34.4 for the Downs component.
Realized habitat (temperature, salinity, water depth, latitude, longitude) of herring by life stage, with NHL subdivided further by spawning component.
| Variable | Source | Area | Life stage | Average of annual medians | s.d. of annual medians | Minimum of first quartile | Maximum of third quartile |
|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 10.6 | 0.9 | 7.5 | 13.1 |
| E | Buchan | NHL | 11.2 | 0.8 | 9.7 | 13.7 | |
| E | Banks | NHL | 11.2 | 1.0 | 8.4 | 15.1 | |
| E | Downs | NHL | 9.3 | 1.2 | 6.1 | 13.5 | |
| E | NS | PML | 5.6 | 1.1 | 2.5 | 8.5 | |
| E | NS | J0 | 13.5 | 1.4 | 9.9 | 17.2 | |
| E | NS | J1 | 4.5 | 0.9 | 0.5 | 7.0 | |
| Salinity | N | Orkney | NHL | 34.9 | 0.0 | 34.8 | 35.1 |
| E | Buchan | NHL | 35.0 | 0.1 | 34.5 | 35.1 | |
| E | Banks | NHL | 34.9 | 0.1 | 34.4 | 35.1 | |
| E | Downs | NHL | 34.4 | 0.2 | 32.6 | 34.9 | |
| E | NS | PML | 33.9 | 0.6 | 32.1 | 35.0 | |
| E | NS | J0 | 33.8 | 0.4 | 32.8 | 34.9 | |
| E | NS | J1 | 33.7 | 0.6 | 31.6 | 35.0 | |
| Depth (m) | E | NS | PML | 52 | 17 | 9 | 105 |
| S | NS | J0 | 38 | 5 | 17 | 103 | |
| S | NS | J1 | 36 | 9 | 13 | 64 | |
| Latitude | E | NS | PML | 54.5 | 1.6 | 51.1 | 58.3 |
| E | NS | J0 | 54.5 | 1.0 | 52.9 | 57.0 | |
| E | NS | J1 | 54.3 | 0.6 | 51.9 | 57.6 | |
| Longitude | E | NS | PML | 1.3 | 2.0 | −3.8 | 6.8 |
| E | NS | J0 | 6.2 | 0.7 | 0.3 | 7.6 | |
| E | NS | J1 | 4.2 | 2.0 | −2.8 | 7.7 |
| Variable | Source | Area | Life stage | Average of annual medians | s.d. of annual medians | Minimum of first quartile | Maximum of third quartile |
|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 10.6 | 0.9 | 7.5 | 13.1 |
| E | Buchan | NHL | 11.2 | 0.8 | 9.7 | 13.7 | |
| E | Banks | NHL | 11.2 | 1.0 | 8.4 | 15.1 | |
| E | Downs | NHL | 9.3 | 1.2 | 6.1 | 13.5 | |
| E | NS | PML | 5.6 | 1.1 | 2.5 | 8.5 | |
| E | NS | J0 | 13.5 | 1.4 | 9.9 | 17.2 | |
| E | NS | J1 | 4.5 | 0.9 | 0.5 | 7.0 | |
| Salinity | N | Orkney | NHL | 34.9 | 0.0 | 34.8 | 35.1 |
| E | Buchan | NHL | 35.0 | 0.1 | 34.5 | 35.1 | |
| E | Banks | NHL | 34.9 | 0.1 | 34.4 | 35.1 | |
| E | Downs | NHL | 34.4 | 0.2 | 32.6 | 34.9 | |
| E | NS | PML | 33.9 | 0.6 | 32.1 | 35.0 | |
| E | NS | J0 | 33.8 | 0.4 | 32.8 | 34.9 | |
| E | NS | J1 | 33.7 | 0.6 | 31.6 | 35.0 | |
| Depth (m) | E | NS | PML | 52 | 17 | 9 | 105 |
| S | NS | J0 | 38 | 5 | 17 | 103 | |
| S | NS | J1 | 36 | 9 | 13 | 64 | |
| Latitude | E | NS | PML | 54.5 | 1.6 | 51.1 | 58.3 |
| E | NS | J0 | 54.5 | 1.0 | 52.9 | 57.0 | |
| E | NS | J1 | 54.3 | 0.6 | 51.9 | 57.6 | |
| Longitude | E | NS | PML | 1.3 | 2.0 | −3.8 | 6.8 |
| E | NS | J0 | 6.2 | 0.7 | 0.3 | 7.6 | |
| E | NS | J1 | 4.2 | 2.0 | −2.8 | 7.7 |
Environmental data derived from E (ECOSMO), N (NORWECOM; only for Orkney spawning ground), or S (survey). The area denotes the spawning area or the whole North Sea (NS). Average and standard deviation (s.d.) of the annual distribution medians, and the minimum of the first quartile and the maximum of the third quartile distribution values. The life stage abbreviation is listed in Table 1.
Realized habitat (temperature, salinity, water depth, latitude, longitude) of herring by life stage, with NHL subdivided further by spawning component.
| Variable | Source | Area | Life stage | Average of annual medians | s.d. of annual medians | Minimum of first quartile | Maximum of third quartile |
|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 10.6 | 0.9 | 7.5 | 13.1 |
| E | Buchan | NHL | 11.2 | 0.8 | 9.7 | 13.7 | |
| E | Banks | NHL | 11.2 | 1.0 | 8.4 | 15.1 | |
| E | Downs | NHL | 9.3 | 1.2 | 6.1 | 13.5 | |
| E | NS | PML | 5.6 | 1.1 | 2.5 | 8.5 | |
| E | NS | J0 | 13.5 | 1.4 | 9.9 | 17.2 | |
| E | NS | J1 | 4.5 | 0.9 | 0.5 | 7.0 | |
| Salinity | N | Orkney | NHL | 34.9 | 0.0 | 34.8 | 35.1 |
| E | Buchan | NHL | 35.0 | 0.1 | 34.5 | 35.1 | |
| E | Banks | NHL | 34.9 | 0.1 | 34.4 | 35.1 | |
| E | Downs | NHL | 34.4 | 0.2 | 32.6 | 34.9 | |
| E | NS | PML | 33.9 | 0.6 | 32.1 | 35.0 | |
| E | NS | J0 | 33.8 | 0.4 | 32.8 | 34.9 | |
| E | NS | J1 | 33.7 | 0.6 | 31.6 | 35.0 | |
| Depth (m) | E | NS | PML | 52 | 17 | 9 | 105 |
| S | NS | J0 | 38 | 5 | 17 | 103 | |
| S | NS | J1 | 36 | 9 | 13 | 64 | |
| Latitude | E | NS | PML | 54.5 | 1.6 | 51.1 | 58.3 |
| E | NS | J0 | 54.5 | 1.0 | 52.9 | 57.0 | |
| E | NS | J1 | 54.3 | 0.6 | 51.9 | 57.6 | |
| Longitude | E | NS | PML | 1.3 | 2.0 | −3.8 | 6.8 |
| E | NS | J0 | 6.2 | 0.7 | 0.3 | 7.6 | |
| E | NS | J1 | 4.2 | 2.0 | −2.8 | 7.7 |
| Variable | Source | Area | Life stage | Average of annual medians | s.d. of annual medians | Minimum of first quartile | Maximum of third quartile |
|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 10.6 | 0.9 | 7.5 | 13.1 |
| E | Buchan | NHL | 11.2 | 0.8 | 9.7 | 13.7 | |
| E | Banks | NHL | 11.2 | 1.0 | 8.4 | 15.1 | |
| E | Downs | NHL | 9.3 | 1.2 | 6.1 | 13.5 | |
| E | NS | PML | 5.6 | 1.1 | 2.5 | 8.5 | |
| E | NS | J0 | 13.5 | 1.4 | 9.9 | 17.2 | |
| E | NS | J1 | 4.5 | 0.9 | 0.5 | 7.0 | |
| Salinity | N | Orkney | NHL | 34.9 | 0.0 | 34.8 | 35.1 |
| E | Buchan | NHL | 35.0 | 0.1 | 34.5 | 35.1 | |
| E | Banks | NHL | 34.9 | 0.1 | 34.4 | 35.1 | |
| E | Downs | NHL | 34.4 | 0.2 | 32.6 | 34.9 | |
| E | NS | PML | 33.9 | 0.6 | 32.1 | 35.0 | |
| E | NS | J0 | 33.8 | 0.4 | 32.8 | 34.9 | |
| E | NS | J1 | 33.7 | 0.6 | 31.6 | 35.0 | |
| Depth (m) | E | NS | PML | 52 | 17 | 9 | 105 |
| S | NS | J0 | 38 | 5 | 17 | 103 | |
| S | NS | J1 | 36 | 9 | 13 | 64 | |
| Latitude | E | NS | PML | 54.5 | 1.6 | 51.1 | 58.3 |
| E | NS | J0 | 54.5 | 1.0 | 52.9 | 57.0 | |
| E | NS | J1 | 54.3 | 0.6 | 51.9 | 57.6 | |
| Longitude | E | NS | PML | 1.3 | 2.0 | −3.8 | 6.8 |
| E | NS | J0 | 6.2 | 0.7 | 0.3 | 7.6 | |
| E | NS | J1 | 4.2 | 2.0 | −2.8 | 7.7 |
Environmental data derived from E (ECOSMO), N (NORWECOM; only for Orkney spawning ground), or S (survey). The area denotes the spawning area or the whole North Sea (NS). Average and standard deviation (s.d.) of the annual distribution medians, and the minimum of the first quartile and the maximum of the third quartile distribution values. The life stage abbreviation is listed in Table 1.
The realized temperature of the interquartile range of the PML was 2.5–8.5°C (Figure 3) over the time-series, with an average annual median of 5.6°C (s.d. 1.1°C; Table 3), and the salinities ranged from 32.1 to 35.0, with an average annual median over the time-series of 33.9 (s.d. 0.6; Table 3).
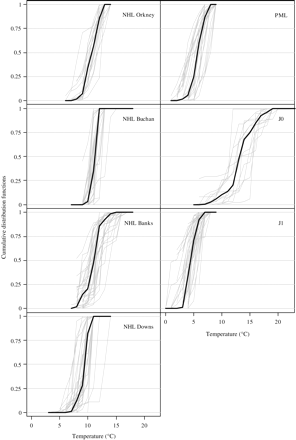
The CDFs of temperature for different life stages of North Sea herring. That of NHL Orkney is based on NORWECOM temperatures, and all others on ECOSMO temperatures.
Juvenile herring aged 0 (J0) were found in warm water in summer (9.9–17.2°C; Figure 3) with an average annual median over the time-series of 13.5°C (s.d. 1.4°C; Table 3). By the following winter, J1 herring are in colder water (0.5–7.0°C; Figure 3) with an average annual median of 4.5°C (s.d. 0.9°C; Table 3). This distribution places them in slightly cooler water than PML at the same time of year. There is little difference between the realized habitats of J0 and J1 for salinity (average annual medians of 33.8 and 33.7, respectively; Table 3). Both classes of juvenile are in shallow water (Table 3) with no clear distinction between the two, though J1 herring are farther west (4.2°W) than J0 (6.2°W; Table 3).
Realized habitats and North Sea trends
The trend in the time-series of realized temperature habitats of the NHL appears to reflect that of the whole North Sea environment (Figure 4). All slope estimates of the linear regression of habitat time-series against environmental time-series are positively correlated. For Buchan NHL and Downs NHL, PML, and J1, the correlations are significant at the 95% level. The herring stages sampled in winter (PML and J1) show the same trend as for the North Sea as a whole, with temperatures slightly increasing over time. The J1 herring are at lower temperatures than the whole North Sea (Figure 4), whereas the J0 herring appear to be in warmer water than the whole North Sea in recent years, with an increasing trend in their realized temperature habitat. This result is at variance with average summer temperatures in the North Sea, which have remained fairly constant over the past decade (Figure 4).
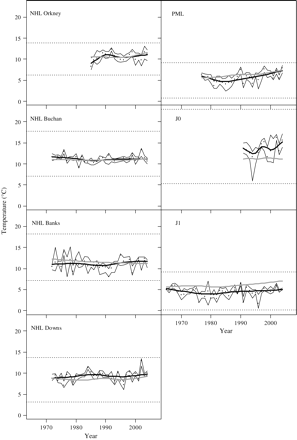
The time-series of realized temperature habitat compared with the environmental trend, shown as the annual median distribution (dots) with a smoother (heavy black line), lower and upper quartiles (thin black lines), and the environmental trend (heavy grey line). Dotted horizontal lines are the minimum and the maximum values observed in any year. NHL Orkney is based on NORWECOM temperatures, and all others on ECOSMO temperatures. Abbreviations are listed in Table 1.
Because of model differences in salinity output, we do not show temporal trends of realized salinity habitats.
The spatial distributions of PML, J0, and J1 herring in the North Sea vary over time (Figure 5), with a gradual shift from north to south. The shift appears to have been strongest for PML, moving from north of 56°N to south of 54°N, and J0 herring, moving from north of 55°N to south of 54°N. The spatial distribution of J1 herring shifted from west of 3°W to east of 5°W, but the interannual variability in this trend is large, shown by the first and the third quartiles. The J1 herring seem to be closer to the coast in the east, because the west–east shift seems to be accompanied by a shift in depth distribution from water deeper than 50 m to water shallower than 40 m.
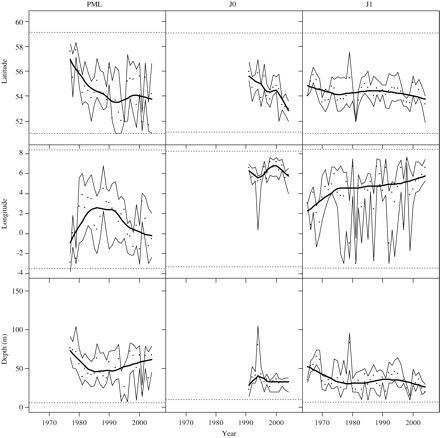
The time-series of the distribution of PML, J0, and J1 herring by latitude (top), longitude (centre), and depth of water column (bottom), showing the realized habitat of the median distribution of the life stage (dots) with a smoother (heavy black line), and lower and upper quartiles (thin black lines). Dotted horizontal lines are the minimum and the maximum values over the time-series.
Discussion
The results here provide an overview of the realized habitats in terms of temperature, salinity, latitude, longitude, and depth of North Sea herring larvae and juveniles (Table 3). Depending on the life stage, changes in the realized habitat could be detected in realized temperature, latitude, longitude, and depth habitat over time (Figures 4 and 5).
As expected, the realized habitats of North Sea herring vary with life stage. The realized habitat of NHL is at the extreme high salinity of the North Sea (>34.4), but the different biophysical models give different descriptions of salinity habitat, so precluding us drawing robust conclusions about the trends in realized salinity habitat of the stock. As they develop, North Sea herring become more prevalent in the areas of lower salinity (a decrease in the average annual median from ∼35.0 to 33.7 from the NHL stages to J1). Several mechanisms control the distribution, and hence the habitat, of herring larvae and juveniles. NHL are located close to the natal spawning grounds that have substrata suitable for successful egg deposition and development (Geffen, 2009); their habitat is chosen for them by the spawners. PML undertake active vertical migration with little horizontal movement, so can be considered virtually as passively drifting larvae (Dickey-Collas et al., 2009). Juvenile herring can shoal, swim actively, and respond to gradients, so to some extent at least can choose their environment.
It is important to note that the North Sea herring stock is not a single, homogenous, well-mixed entity; the individual components have unique population dynamics and their contribution to the overall stock varies appreciably throughout time (Cushing and Bridger, 1966; Schmidt et al., 2009; Payne, 2010). Changes in the relative composition of the overall stock will change the number of larvae released at each spawning ground and can be expected to influence the spatial distribution of at least PML, and possibly J0 and even J1 herring. The increasingly southern distribution of PML and J0 (Figure 5) may therefore reflect the relative increase in the southern Downs component reported elsewhere (Payne, 2010), rather than an active process of habitat selection.
In the same way that the North Sea herring stock is not homogenous, the North Sea itself displays a high degree of heterogeneity; the realized habitat is closely tied to the spatial distribution of the fish. The more southern distribution of J0 herring is therefore the most likely explanation for the increase in that group's realized temperature habitat (Figure 4), as well as for the lack of correlation between trends in the environment and this particular realized habitat (Table 4). The North Sea generally increases in temperature in summer towards the south, so a more southerly distribution will, on average, increase the average temperature experienced. This example serves as a reminder of the multidimensional nature of the problem and the weakness of the approach; other, usually “unseen”, variables can often be more influential than the single parameter being evaluated.
Analysis of trends in realized habitat of early life stages with trends in the environment.
| Variable | Source | Area | Life stage | Slope estimate | Standard error | t | Pr(>|t|) | Multiple R2 |
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 0.12 | 0.11 | 1.05 | 0.309 | 0.06 |
| E | Buchan | NHL | 0.14 | 0.05 | 2.65 | 0.013 | 0.19 | |
| E | Banks | NHL | 0.08 | 0.09 | 0.91 | 0.368 | 0.03 | |
| E | Downs | NHL | 0.36 | 0.11 | 3.19 | 0.003 | 0.25 | |
| E | North Sea | PML | 0.43 | 0.09 | 4.90 | 0.000 | 0.48 | |
| E | North Sea | J0 | 0.06 | 0.09 | 0.70 | 0.497 | 0.04 | |
| E | North Sea | J1 | 0.29 | 0.10 | 2.80 | 0.008 | 0.17 | |
| Salinity | N | Orkney | NHL | 0.02 | 0.05 | 0.43 | 0.669 | 0.01 |
| E | Buchan | NHL | 0.20 | 0.06 | 3.46 | 0.007 | 0.57 | |
| E | Banks | NHL | 0.28 | 0.08 | 3.37 | 0.002 | 0.29 | |
| E | Downs | NHL | 0.16 | 0.17 | 1.00 | 0.326 | 0.03 | |
| E | North Sea | PML | 0.04 | 0.04 | 1.09 | 0.288 | 0.04 | |
| E | North Sea | J0 | 0.04 | 0.04 | 1.02 | 0.328 | 0.08 | |
| E | North Sea | J1 | 0.11 | 0.03 | 3.12 | 0.003 | 0.20 |
| Variable | Source | Area | Life stage | Slope estimate | Standard error | t | Pr(>|t|) | Multiple R2 |
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 0.12 | 0.11 | 1.05 | 0.309 | 0.06 |
| E | Buchan | NHL | 0.14 | 0.05 | 2.65 | 0.013 | 0.19 | |
| E | Banks | NHL | 0.08 | 0.09 | 0.91 | 0.368 | 0.03 | |
| E | Downs | NHL | 0.36 | 0.11 | 3.19 | 0.003 | 0.25 | |
| E | North Sea | PML | 0.43 | 0.09 | 4.90 | 0.000 | 0.48 | |
| E | North Sea | J0 | 0.06 | 0.09 | 0.70 | 0.497 | 0.04 | |
| E | North Sea | J1 | 0.29 | 0.10 | 2.80 | 0.008 | 0.17 | |
| Salinity | N | Orkney | NHL | 0.02 | 0.05 | 0.43 | 0.669 | 0.01 |
| E | Buchan | NHL | 0.20 | 0.06 | 3.46 | 0.007 | 0.57 | |
| E | Banks | NHL | 0.28 | 0.08 | 3.37 | 0.002 | 0.29 | |
| E | Downs | NHL | 0.16 | 0.17 | 1.00 | 0.326 | 0.03 | |
| E | North Sea | PML | 0.04 | 0.04 | 1.09 | 0.288 | 0.04 | |
| E | North Sea | J0 | 0.04 | 0.04 | 1.02 | 0.328 | 0.08 | |
| E | North Sea | J1 | 0.11 | 0.03 | 3.12 | 0.003 | 0.20 |
Linear regression parameters: slope estimate, standard error, t-value, probability of t-value, and multiple R2. Emboldened p-values are significant at the >95% confidence level. Model sources ECOSMO (E) and NORWECOM (N). Life stages described in Table 1.
Analysis of trends in realized habitat of early life stages with trends in the environment.
| Variable | Source | Area | Life stage | Slope estimate | Standard error | t | Pr(>|t|) | Multiple R2 |
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 0.12 | 0.11 | 1.05 | 0.309 | 0.06 |
| E | Buchan | NHL | 0.14 | 0.05 | 2.65 | 0.013 | 0.19 | |
| E | Banks | NHL | 0.08 | 0.09 | 0.91 | 0.368 | 0.03 | |
| E | Downs | NHL | 0.36 | 0.11 | 3.19 | 0.003 | 0.25 | |
| E | North Sea | PML | 0.43 | 0.09 | 4.90 | 0.000 | 0.48 | |
| E | North Sea | J0 | 0.06 | 0.09 | 0.70 | 0.497 | 0.04 | |
| E | North Sea | J1 | 0.29 | 0.10 | 2.80 | 0.008 | 0.17 | |
| Salinity | N | Orkney | NHL | 0.02 | 0.05 | 0.43 | 0.669 | 0.01 |
| E | Buchan | NHL | 0.20 | 0.06 | 3.46 | 0.007 | 0.57 | |
| E | Banks | NHL | 0.28 | 0.08 | 3.37 | 0.002 | 0.29 | |
| E | Downs | NHL | 0.16 | 0.17 | 1.00 | 0.326 | 0.03 | |
| E | North Sea | PML | 0.04 | 0.04 | 1.09 | 0.288 | 0.04 | |
| E | North Sea | J0 | 0.04 | 0.04 | 1.02 | 0.328 | 0.08 | |
| E | North Sea | J1 | 0.11 | 0.03 | 3.12 | 0.003 | 0.20 |
| Variable | Source | Area | Life stage | Slope estimate | Standard error | t | Pr(>|t|) | Multiple R2 |
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | N | Orkney | NHL | 0.12 | 0.11 | 1.05 | 0.309 | 0.06 |
| E | Buchan | NHL | 0.14 | 0.05 | 2.65 | 0.013 | 0.19 | |
| E | Banks | NHL | 0.08 | 0.09 | 0.91 | 0.368 | 0.03 | |
| E | Downs | NHL | 0.36 | 0.11 | 3.19 | 0.003 | 0.25 | |
| E | North Sea | PML | 0.43 | 0.09 | 4.90 | 0.000 | 0.48 | |
| E | North Sea | J0 | 0.06 | 0.09 | 0.70 | 0.497 | 0.04 | |
| E | North Sea | J1 | 0.29 | 0.10 | 2.80 | 0.008 | 0.17 | |
| Salinity | N | Orkney | NHL | 0.02 | 0.05 | 0.43 | 0.669 | 0.01 |
| E | Buchan | NHL | 0.20 | 0.06 | 3.46 | 0.007 | 0.57 | |
| E | Banks | NHL | 0.28 | 0.08 | 3.37 | 0.002 | 0.29 | |
| E | Downs | NHL | 0.16 | 0.17 | 1.00 | 0.326 | 0.03 | |
| E | North Sea | PML | 0.04 | 0.04 | 1.09 | 0.288 | 0.04 | |
| E | North Sea | J0 | 0.04 | 0.04 | 1.02 | 0.328 | 0.08 | |
| E | North Sea | J1 | 0.11 | 0.03 | 3.12 | 0.003 | 0.20 |
Linear regression parameters: slope estimate, standard error, t-value, probability of t-value, and multiple R2. Emboldened p-values are significant at the >95% confidence level. Model sources ECOSMO (E) and NORWECOM (N). Life stages described in Table 1.
The distributions in temperature of herring larvae (NHL and PML) closely follow environmental trends (Figure 4), suggesting that the changes in North Sea temperature fall within the potential temperature habitat for herring and that the habitat of these life stages is likely indifferent to the changes in temperature. This is not surprising, because these fish cannot actively select their habitat, so their distribution would be expected to reflect changes in their environment.
Climate models predict a future increase in air and water temperature (Drinkwater, 2010). The PML show a slight increase in the realized temperature habitat over the past 2–3 decades. Surrounding North Sea temperatures appear to have increased slightly during winters. Additionally, the realized temperature habitat of PML and J1 herring in winter appears to be in waters slightly (0–2°C) cooler than the average of the surrounding North Sea. If these trends are the result of the realized habitat of herring being constrained to the areas of lower temperature, then further warming in future may cause habitat limitation. However, they could equally be the result of some other constraining variable controlling the distribution of herring, in which case warming may not have a significant effect.
North Sea herring use a subset of the North Sea environment as realized habitat. Shallower waters experience greater variability in temperature over the year than deeper areas of the North Sea. J1 herring are found in water that is 1–3°C cooler than the average North Sea (Figure 4), and over the past three decades they have shifted more to the east, towards shallower water closer to the coast (Figure 5). One possible explanation is that the larger J1 herring are more able to control their location and response to the environment. It could be that the latter try to reduce metabolic energy costs by choosing colder water during winter, when they are not feeding. If so, it would suggest that temperature, although not constraining distribution, would have the potential to impact the dynamics of herring.
The temperature difference between the Downs and the other spawning locations is ∼2°C. This difference impacts on the physiological requirements of the larvae and the development of meristic characteristics (Heath et al., 1997). Herring larvae can grow and survive in temperatures from 1 to at least 17°C (Oeberst et al., 2009) and in the North Atlantic the potential temperature habitat of herring ranges between a mean temperature of 3 and 15°C (Brunel and Dickey-Collas, 2010), placing the North Sea in the middle of the range. Drifting after hatching, the larvae are transported east towards cooler, less-saline habitat (Figure 5), and it is during this phase that recruitment strength is determined (Nash and Dickey-Collas, 2005; Payne et al., 2009).
The survey data used in this study are derived from ICES coordinated surveys, which are quality-assured and executed using standard protocols. Therefore, like many other studies, we have assumed that the data are suitable for the analysis we have carried out (Simmonds, 2009). The habitat analysis is based on environmental data derived from biophysical models, but it can be argued that such an analysis should use empirical observations only. However, complete, consistent, quality-assured datasets of the appropriate length and resolution do not exist. If we had put together a collection of best available data into a concatenated time-series, apart from gaps there would also be problems in understanding whether changes in variability were from changes in sampling (and quality control) or the actual environmental signal. It is for that reason that we used two series of available modelled datasets and compared the results.
The ECOSMO and the NORWECOM models have been scrutinized during several studies of the North Sea (Skogen and Moll, 2000; Daewel et al., 2008), and the NORWECOM output has been validated extensively by comparison with field data (Søiland and Skogen, 2000). The use of ECOSMO or NORWECOM to define the physical environment produced similar datasets and descriptions of realized habitats for temperature, but not for salinity. The salinity variance can be attributed to a number of processes affecting the models, i.e. the differences in model setup (ECOSMO for the North Sea and the Baltic Sea, NORWECOM for the Nordic Sea and North Sea), and the consequent differences in North Sea salinity boundary conditions, to differences in freshwater run-off forcing, as well as differences in evaporation and precipitation (Schrum and Backhaus, 1999; Schrum et al., 2003). In general, the accuracy of model data tends to be less close to boundaries. However, ECOSMO is more appropriate for considering the salinity habitat of juvenile herring close to the Skagerrak, because it covers jointly the North and the Baltic Seas.
Similar methods have been used to determine the distribution probabilities of an organism with respect to environmental and geographic covariates and to account for the sampling bias in other studies (e.g. Tomkiewicz et al., 1998; Castonguay et al., 1999). Most methods with the potential to describe habitats fail to account properly for sampling bias, e.g. single parameter quotients (Bernal et al., 2007; Ibaibarriaga et al., 2007). The method employed here corrects for the effect of sampling, so can characterize the realized habitat, but the limiting factor appears to be the ability of the models to describe the environmental variable over time and space adequately, i.e. the quality of the information about the environmental variables, rather than the method used.
Acknowledgements
The study was funded by the EU FP6 research projects RECLAIM and UNCOVER. Corinna Schrum, Irina Alekseeva, and Morten Skogen are acknowledged for providing the biophysical model data and discussing the results, and Pierre Petitgas, Adriaan Rijnsdorp, Niels Hintzen, and Jan Jaap Poos for advice. The comments and suggestions of four anonymous reviewers and editor Audrey Geffen were greatly appreciated and helped to improve the manuscript.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}