





Abstract
An empirical harvest strategy for an Australian longline fishery was developed and tested using harvest strategy evaluation. The approach is based on catch rate and size composition indices and iteratively drives the population towards a target level of spawning potential ratio. The simulation model assumed traditional age-structured population dynamics. The results demonstrate that the empirical approach represents a promising feedback strategy that becomes more precautionary as data errors and recruitment variance increase. Given the urgent need for formal harvest strategies to replace ad hoc decision-making in data-poor fisheries, the approach offers considerable promise. Independent implementation at local scales across the range of a species is speculated to result in management that is robust to uncertainty about stock structure.Prince, J. D., Dowling, N. A., Davies, C. R., Campbell, R. A., and Kolody, D. S. 2011. A simple cost-effective and scale-less empirical approach to harvest strategies. – ICES Journal of Marine Science, 68: 947–960.
Introduction
The field of modern fisheries reflects its origins in large industrial-scale resources. Much of the current work on harvest policies, decision rules, and their evaluation focuses on relatively large-scale and geographically extensive fisheries and the use of indices of biomass (B) or fishing intensity (F) estimated using stock assessment models (e.g. Sainsbury et al., 2000; Punt et al., 2001). However, assessments may be sensitive to constraining assumptions, and multiple model specifications might be consistent with data, but indicative of different reference points and management implications (Schnute and Richards, 2001). Additionally, many fisheries are data-poor, preventing the application of advanced assessment models. While overfishing intensifies (Mullon et al., 2005), many approaches to address overfishing require data and model structures beyond the capability of most fisheries (Walters and Pearse, 1996; Cochrane, 1999). There is a need for simpler assessment and management procedures based on indicators collected directly from the catch and/or from simple surveys (Johannes, 1998; Berkes et al., 2001; Orensanz et al., 2005; Ostrom, 2007; Prince, 2010). This paper outlines an empirical approach for setting catch quota based on local size-specific catch per unit effort (cpue) for the Australian eastern tuna and billfish fishery (ETBF) and describes a pilot evaluation of the approach for broadbill swordfish (Xiphias gladius).
Froese (2004) suggested that assessments could be based on three size-based indices from catch composition data, Px: (i) the percentage of mature fish in the catch, Pmat, with 100% as target; (ii) the percentage of fish with optimum length in the catch, Popt, with 100% as target; and (iii) the percentage of large fish in the catch, Pmega, with 0% as target, and 30–40% as representative of reasonable stock structure if no upper size limit exists. Cope and Punt (2009) showed that Froese's (2004) values were not always sufficient to ensure protection from overfishing and introduced Pobj (the sum of Pmat, Popt, and Pmega) to distinguish selectivity patterns.
We present a harvest rule for which size-based catch rates are the main driver, and directly index stock depletion in the model used to test the rule. Size composition catch data are decomposed into three classes, small (“recruits”), medium (“prime”), and large (“old”). Indicators consisting of size-based catch rates, and the proportion of old fish are evaluated within a decision-tree framework relative to the stock's predicted size composition at equilibrium around the target levels of prime-sized cpue and spawning per recruit, or spawning potential ratio. For convenience we follow the latter, defining spawning potential ratio (SPR) as the proportion of unfished reproductive potential allowed by harvesting (Walters and Martell, 2004). Within our context of equilibrium modelling, the proportion of lifetime spawning per recruit (Mace and Sissenwine, 1993) equals the proportion of a stock's seasonal spawning potential allowed by harvesting (Walters and Martell, 2004).
This approach both builds on Caddy's (1998, 2002) traffic-light concept and attempts to operationalize and evaluate the utility of Froese's (2004) notion of keeping it simple. Replacement SPR has been recommended by Mace and Sissenwine (1993) as a reference point for defining overfishing; it is based on the premise of stock replacement, and in the absence of a formal stock assessment, taxonomic affiliation and life-history parameters can be used to select preliminary estimates. Our framework also builds on the principle of Mace et al. (1996) that length distribution, or SPR alone, can be ambiguous and an overly optimistic index of stock abundance. Our combined use of cpue and size-based SPR reference points to cross-check simpler cpue trends addresses the weaknesses identified by Cope and Punt (2008) in Froese's (2004) approach.
East coast tuna and billfish fishery
The most valuable species in the ETBF are broadbill swordfish (X. gladius), bigeye tuna (Thunnus obesus), and yellowfin tuna (T. albacares), with an annual landed value of ∼$AUD45 million. Catches generally comprise <1% of western and central Pacific Ocean (WCPO) landings (Campbell, 2007). The ETBF fishes a peripheral pocket of stock in the Coral and Tasman Seas (CTS). The extent of movement and mixing are uncertain (Sibert and Hampton, 2002; Campbell and Hobday, 2003), with recent studies emphasizing site fidelity within the CTS. The relevance of trends described by WCPO stock assessments to smaller CTS stocks is debatable.
The Australian Government policy “Securing our Fishing Future” aims to stop overfishing and rebuild overfished fish stocks (Australian Government, 2007) via harvest policies for Commonwealth fisheries (McDonald, 2005) consistent with established guidelines (Australian Government, 2007). Standard model estimates of biomass and fishing mortality for the ETBF are sensitive to assumptions on stock structure. Trends measured for the larger WCPO stocks, which would dominate modelled dynamics, may not be indicative of smaller, potentially weakly linked CTS stocks.
We outline an empirical harvest strategy developed for the ETBF using size-based catch rates as local indicators of stock status (Davies et al., 2008). The assumption is that the Australian fleet will be managed in response to impacts experienced by the stocks they fish, whether locally or by accumulation over the broader WCPO. Management based on the size composition and catch rates of locally caught fish is considered precautionary, but if there are linkages with more-heavily fished WCPO stocks, it is likely that local trends in size-based catch rates, and hence the recommended biological catch (RBC) will be affected irrespective of whether the Australian fleet is primarily responsible for the impact. Depending on how global catches are regulated, fishing pressure may not be reduced significantly across a broader WCPO stock. Conversely, if CTS stocks are relatively isolated, good management of the ETBF would not depend on international agreement.
We describe the hierarchical decision tree incorporating size-based catch rates to assess the CTS stocks relative to equilibrium catch rates associated with target levels of SPR and provide the results of an initial evaluation via simulations of a unit stock. Despite uncertainty regarding the validity of the unit-stock assumption, the study provides a pragmatic first step in evaluating the empirically based decision tree. If the rule performs poorly in the unit-stock context, it would be unlikely to be useful when the connection between local management and the population response is less direct.
The decision tree
Assessment of SPR
The decision tree (Figure 1) uses the logic of virtual population analysis (Gulland, 1969) and size-based catch rates to assess catch composition data relative to those expected if the stock was at some equilibrium target level of SPR. The RBC is calculated based on a target catch rate, then decreased incrementally if SPR is inferred from size-based catch rates to be below the target level and increased if the SPR is inferred to be above the target. Estimates of sustainable catch emerge indirectly as the catch level at which the stock stabilizes around the target level of SPR.
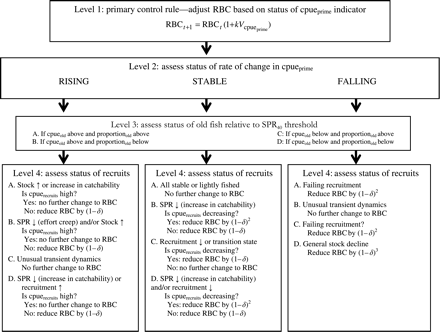
Flowchart diagram of the four levels of the decision tree. At Level 1, the primary control rule calculates the new RBC based on the catch rate of the prime-sized fish (cpueprime). Note that δx, where x = 1, 2, or 3, is the reduction factor, expressed as the remaining proportion of the total. For example, a 10% reduction from an action where the RBC is reduced by (1 − δx) would imply that δ = 0.9 and x = 1.
A reference level SPR is translated into a size or age structure with a model constructed incorporating size-at-age distributions, age- or size-fecundity ogives, and an estimate of natural mortality. This is used to estimate the size-based catch rates corresponding to the reference or the target SPR level. Expected values of cpue for the unexploited stock and target rates are calculated using an operating model that assumes a known stock–recruitment function. Under the assumption of equilibrium, in the absence of recruitment trends, and assuming that steepness equals 1, SPR can be used to infer relative abundance levels corresponding to the equilibrium SPR reference level. Population size structure indicates an approximate level of SPR that can be used to infer a depletion level (O'Farrell and Botsford, 2005). However, the population may be far from equilibrium, and size composition may be much affected by recent recruitment trends, so other forms of data could ultimately be useful in calibrating the size-based catch rates. The ETBF has catch rate and size data from the 1950s, when CTS stocks were relatively lightly exploited. If small spatial scale and localized nature of populations confer uncertainty on stock assessments, marine protected areas could provide proxies for virgin size-based catch rates (Wilson et al., 2010), although these would be of little use for highly migratory species. Otherwise, the decision tree would need to be calibrated using SPR models and expert judgement on current depletion. The impact of imprecision and misspecification when setting reference levels of SPR is tested below.
The hierarchical decision tree yields a preliminary estimate of the incremental change in catch with respect to the previous RBC based on the cpue trend of the prime size group. This initial estimate of the incremental change in catch is adjusted according to a series of decision branches selected initially on whether the prime size cpue trend is rising, stable, or falling. Secondary information is derived based on trends in the largest/oldest component of the sampled population and, if the interpretation of trends remains ambiguous, from trends in the smallest fish. The two extremes of size composition provide information that may not be apparent from the aggregated cpue trend. For example, a steady increase in fishing efficiency could maintain or increase aggregated and primary cpue, masking underlying population decline and escalating fishing pressure. The primary cpue-based decision rule would respond by maintaining or increasing catch levels. However, if the proportion and/or the catch rate of the largest fish is declining, the decision tree will act to decrease the RBC incrementally.
Information used
The decision tree uses four size-based indicators: cpue of each of the three size classes (cpueprime, cpueold, and cpuerecruits) and the proportion of old fish (proportionold). ETBF catch is reported as individual fish weights (Campbell et al., 2007), which are divided arbitrarily into small (recruits; <50 kg), medium (prime; 50–150 kg), and large (old; >150 kg) categories. The term recruits refers to the smallest part of the size distribution and may include several of the youngest age classes.
The target reference point for the cpueold and proportionold indicators are set as 40% of spawning potential ratio (SPR40), a BMSY proxy as per the Australian Government policy. The equilibrium age–size-structured model is used to calculate the size composition for the population corresponding to the management target level of SPR40, and hence the cpueold and proportionold targets by size category.
The simulated swordfish population is considered to be at the virgin level in the first year of the simulations, so the initial cpueprime is an index of the unexploited prime-sized abundance. The simulated population is seeded via the specification of a fixed initial population size and as such is not conditioned on data. The target value for cpueprime, cpuetarg, used in the primary control rule below, was taken to be 60% of its initial (specified) level, as a proxy approximation of BMSY ∼ 0.40 B0, assuming that the initial level equates to fishing a virgin stock.
The decision tree has four so-called levels (Figure 1, Table 1), described below.
Listing of the parameters used in the decision tree.
| Decision level | Parameter | Suggested value |
|---|---|---|
| 1 | Number of years over which the slope of cpueprime is calculated | 5 years |
| Target value for cpueprime when calculating slope-to-target | cpueprime at MSY = 0.6 × initial cpueprime | |
| Number of years to rebuild to target value | 5 years | |
| Feedback gain/responsiveness factor, k | 0.7 | |
| 2 | Bound on the percentage annual change in cpueprime to define stability in this indicator (note that the change is relative to the mean value of cpueprime over the previous y years—see below) | 5% per year |
| Number of years over which mean cpueprime is calculated | 5 years | |
| 3 | Target value for cpueold | Cpueold corresponding to that at SPR40 |
| Target value for proportionold | Proportionold corresponding to that at SPR40 | |
| 4 | Value of cpuerecruits to define high recruitment | 0.7 initial cpuerecruits |
| Decrease in cpuerecruits to define declining recruitment | 10% per yeara | |
| Reduction factor on RBC | 10% | |
| Number of years over which mean cpuerecruits is calculated | 5 years |
| Decision level | Parameter | Suggested value |
|---|---|---|
| 1 | Number of years over which the slope of cpueprime is calculated | 5 years |
| Target value for cpueprime when calculating slope-to-target | cpueprime at MSY = 0.6 × initial cpueprime | |
| Number of years to rebuild to target value | 5 years | |
| Feedback gain/responsiveness factor, k | 0.7 | |
| 2 | Bound on the percentage annual change in cpueprime to define stability in this indicator (note that the change is relative to the mean value of cpueprime over the previous y years—see below) | 5% per year |
| Number of years over which mean cpueprime is calculated | 5 years | |
| 3 | Target value for cpueold | Cpueold corresponding to that at SPR40 |
| Target value for proportionold | Proportionold corresponding to that at SPR40 | |
| 4 | Value of cpuerecruits to define high recruitment | 0.7 initial cpuerecruits |
| Decrease in cpuerecruits to define declining recruitment | 10% per yeara | |
| Reduction factor on RBC | 10% | |
| Number of years over which mean cpuerecruits is calculated | 5 years |
aChange defined in a similar manner as for the Level 2 parameter.
Listing of the parameters used in the decision tree.
| Decision level | Parameter | Suggested value |
|---|---|---|
| 1 | Number of years over which the slope of cpueprime is calculated | 5 years |
| Target value for cpueprime when calculating slope-to-target | cpueprime at MSY = 0.6 × initial cpueprime | |
| Number of years to rebuild to target value | 5 years | |
| Feedback gain/responsiveness factor, k | 0.7 | |
| 2 | Bound on the percentage annual change in cpueprime to define stability in this indicator (note that the change is relative to the mean value of cpueprime over the previous y years—see below) | 5% per year |
| Number of years over which mean cpueprime is calculated | 5 years | |
| 3 | Target value for cpueold | Cpueold corresponding to that at SPR40 |
| Target value for proportionold | Proportionold corresponding to that at SPR40 | |
| 4 | Value of cpuerecruits to define high recruitment | 0.7 initial cpuerecruits |
| Decrease in cpuerecruits to define declining recruitment | 10% per yeara | |
| Reduction factor on RBC | 10% | |
| Number of years over which mean cpuerecruits is calculated | 5 years |
| Decision level | Parameter | Suggested value |
|---|---|---|
| 1 | Number of years over which the slope of cpueprime is calculated | 5 years |
| Target value for cpueprime when calculating slope-to-target | cpueprime at MSY = 0.6 × initial cpueprime | |
| Number of years to rebuild to target value | 5 years | |
| Feedback gain/responsiveness factor, k | 0.7 | |
| 2 | Bound on the percentage annual change in cpueprime to define stability in this indicator (note that the change is relative to the mean value of cpueprime over the previous y years—see below) | 5% per year |
| Number of years over which mean cpueprime is calculated | 5 years | |
| 3 | Target value for cpueold | Cpueold corresponding to that at SPR40 |
| Target value for proportionold | Proportionold corresponding to that at SPR40 | |
| 4 | Value of cpuerecruits to define high recruitment | 0.7 initial cpuerecruits |
| Decrease in cpuerecruits to define declining recruitment | 10% per yeara | |
| Reduction factor on RBC | 10% | |
| Number of years over which mean cpuerecruits is calculated | 5 years |
aChange defined in a similar manner as for the Level 2 parameter.
Level 1: primary control rule—adjust RBC based on cpueprime
The Level 1 control rule updates the RBC according to the distance of cpueprime from its target and the time-frame over which the target is to be reached, both of which form an objective that determines a path on which catches are adjusted to remain.
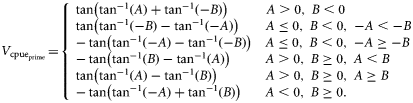
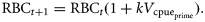
![Conceptual examples of how the slope-to-target term is derived. θs is the angle from the horizontal to the linear regression of the cpue over the previous x years, and θt the angle from the horizontal to the linear path from the current (most recent annual) cpue to the target cpue in y years. In this instance, the slope-to-target = tan(θs + θt) = tan[tan−1(A) + tan−1(B)]. More generally, the slope-to-target is the overall difference between the slope of the cpue trend over the previous x years and the slope from the current cpue to the target cpue achieved in y years.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/icesjms/68/5/10.1093_icesjms_fsr029/1/m_fsr02902.gif?Expires=1716339483&Signature=paZ-kdvgZVdYIsErZccB7I3Ihi~ZyjgsqEpMQZ3vsmwRcxbsy5C~ieATJLbjb0yTAMydya6w02U8uPuV3HNxPvd9~JFe5LMZvDbvBBF8UTiRM5hIfhwkLe1SIVMso1VXrGaLGoP9gkgjNBsukdj7V8CmKAoN27XziUwd7OBgAEgqgp9OrEb6UdlUM-BNMHu8hsjYYHUZQgNQAtHB1GY6YRfwymkBczKXffwPSro5TNfNStsxxPZl-nFkS3~5jAz~6ACYiyOMipYAzcz~kWgNaVmgFS0Mk1sFOKNneg95nIYJH-HRKu72Zs6oojcdCwfrz7LMRjSLacqHfVMnBE7viw__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Conceptual examples of how the slope-to-target term is derived. θs is the angle from the horizontal to the linear regression of the cpue over the previous x years, and θt the angle from the horizontal to the linear path from the current (most recent annual) cpue to the target cpue in y years. In this instance, the slope-to-target = tan(θs + θt) = tan[tan−1(A) + tan−1(B)]. More generally, the slope-to-target is the overall difference between the slope of the cpue trend over the previous x years and the slope from the current cpue to the target cpue achieved in y years.
Level 2: assess status of the rate of change in cpueprime
Cpueprime is categorized as rising, stable, or falling, where stability is defined as the annual rate of change, defined as the slope of a linear regression, being within a given limit (in this case, 5% of the average cpueprime over 5 years). This second level of the decision tree uses the simple trend in cpueprime to determine which of its three main limbs will be used.
Level 3: assess status of old fish relative to SPR40 threshold
Here, it is determined whether cpueold and proportionold values are above or below the target (Figure 1). The four possible outcomes have different interpretations depending on how cpueprime was categorized at Level 2. Both cpue and the proportion of old fish are used because their potential contrast is informative, increasing the discriminatory power of the assessment. For example, if cpueprime is rising and cpueold and proportionold are both above the target, either the stock or the catchability is increasing (Figure 1). However, if cpueold is above the target and proportionold below the target, a reduction in spawner biomass-per-recruit and/or a stock increase would be suggested. Interpretation is finalized by considering the status of the smallest fish (recruits) in Level 4 of the decision tree, described below.
Level 4: assess status of recruits
For some combinations of the status of cpueprime, cpueold and proportionold relative to their respective targets, the trend in cpuerecruits is considered. As changes in proportionold may be produced by a change in the rates of recruitment rather than a change in the actual absolute number of old fish, cpuerecruits provides a final test for distinguishing between otherwise ambiguous possibilities. In certain cases, the RBC is adjusted according to whether cpuerecruits is considered to be greater than a reference level of 70% of its unfished level (high), or if the slope of a linear regression of cpuerecruits over the past 5 years indicates a decline of >10% per year.
For sensitivity analyses where Level 4 is omitted from the decision tree, the status of cpuerecruits is considered to be high and not decreasing. Based on the outcomes from Levels 2–4, the initial RBC from the primary control rule may be reduced by (1 − δ)x, where δ = 0.1 and x = 1, 2, or 3 (Figure 1).
Life-history and design parameters
The simulation model is a simple age-structured population model (see the Supplementary material) developed by Basson and Dowling (2007). Quarterly time-steps account for seasonal spawning and recruitment, the latter occurring in the first two quarters of the year and natural and fishing mortality at the start of each quarter. The plus-group age was 29 years. The model population has no spatial structure, is harvested by a single fleet, and represents a single stock with no movement dynamics.
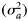 with (annual) age ã, based on Young and Drake (2004):
with (annual) age ã, based on Young and Drake (2004): 
Swordfish population and fishery parameters based on Kolody et al. (2006) and decision-tree implementation criteria used for the harvest strategy evaluation reference scenario.
| Population parameter | Base value | Reference | Definition and/or rationale |
|---|---|---|---|
| Steepness, h | 0.65 | Basson and Dowling (2007) | Ratio between recruitment (R) when spawning biomass is at 20% of unexploited spawning biomass (B0), and recruitment at the unexploited spawning biomass i.e. h = R(0.2 B0)/R(B0) |
| von Bertalanffy growth parameter, k | Male 0.1096, female 0.0815 | Young and Drake (2004) | Averaged so that there is no sexual dimorphism (in the reference case) |
| Natural mortality, M | 0.2 for all ages | ||
| Age at first maturity, tm | 9 | Young and Drake (2004) | |
| Standard deviation of the logarithm of the variability around the stock–recruitment curve, σR | 0.6 | Log(variation in year-class strength) | |
| Initial recruitment, InitR | 200 000 fish | ||
| Fishery parameter | |||
| Catchability, q | 1 × 10−5 | Yield magnitudes comparable with those historically reported in units of numbers per 1000 hooks | |
| Standard deviation of the logarithm of the variability in catch rate, σC | 0.2 | ||
| Size ranges corresponding to recruits, prime, and old fish | Recruits <50 kg, prime 50–150 kg, old >150 kg | Obtained from discussions with ETBF industry members | |
| Decision-tree implementation | |||
| Number of years after which to invoke decision tree | 5 | ||
| Annual fishing mortality before invoking decision tree | 3 FMSY | High level of initial depletion to test the ability of the harvest strategy to rebuild the stock | |
| Frequency of RBC recalculation using decision tree | Annually | ||
| Sampling of catch | |||
| Perfect | 100% | ||
| Moderate, reflecting real-life sampling proportions | 70% | ||
| Poor | 2% | ||
| Population parameter | Base value | Reference | Definition and/or rationale |
|---|---|---|---|
| Steepness, h | 0.65 | Basson and Dowling (2007) | Ratio between recruitment (R) when spawning biomass is at 20% of unexploited spawning biomass (B0), and recruitment at the unexploited spawning biomass i.e. h = R(0.2 B0)/R(B0) |
| von Bertalanffy growth parameter, k | Male 0.1096, female 0.0815 | Young and Drake (2004) | Averaged so that there is no sexual dimorphism (in the reference case) |
| Natural mortality, M | 0.2 for all ages | ||
| Age at first maturity, tm | 9 | Young and Drake (2004) | |
| Standard deviation of the logarithm of the variability around the stock–recruitment curve, σR | 0.6 | Log(variation in year-class strength) | |
| Initial recruitment, InitR | 200 000 fish | ||
| Fishery parameter | |||
| Catchability, q | 1 × 10−5 | Yield magnitudes comparable with those historically reported in units of numbers per 1000 hooks | |
| Standard deviation of the logarithm of the variability in catch rate, σC | 0.2 | ||
| Size ranges corresponding to recruits, prime, and old fish | Recruits <50 kg, prime 50–150 kg, old >150 kg | Obtained from discussions with ETBF industry members | |
| Decision-tree implementation | |||
| Number of years after which to invoke decision tree | 5 | ||
| Annual fishing mortality before invoking decision tree | 3 FMSY | High level of initial depletion to test the ability of the harvest strategy to rebuild the stock | |
| Frequency of RBC recalculation using decision tree | Annually | ||
| Sampling of catch | |||
| Perfect | 100% | ||
| Moderate, reflecting real-life sampling proportions | 70% | ||
| Poor | 2% | ||
Swordfish population and fishery parameters based on Kolody et al. (2006) and decision-tree implementation criteria used for the harvest strategy evaluation reference scenario.
| Population parameter | Base value | Reference | Definition and/or rationale |
|---|---|---|---|
| Steepness, h | 0.65 | Basson and Dowling (2007) | Ratio between recruitment (R) when spawning biomass is at 20% of unexploited spawning biomass (B0), and recruitment at the unexploited spawning biomass i.e. h = R(0.2 B0)/R(B0) |
| von Bertalanffy growth parameter, k | Male 0.1096, female 0.0815 | Young and Drake (2004) | Averaged so that there is no sexual dimorphism (in the reference case) |
| Natural mortality, M | 0.2 for all ages | ||
| Age at first maturity, tm | 9 | Young and Drake (2004) | |
| Standard deviation of the logarithm of the variability around the stock–recruitment curve, σR | 0.6 | Log(variation in year-class strength) | |
| Initial recruitment, InitR | 200 000 fish | ||
| Fishery parameter | |||
| Catchability, q | 1 × 10−5 | Yield magnitudes comparable with those historically reported in units of numbers per 1000 hooks | |
| Standard deviation of the logarithm of the variability in catch rate, σC | 0.2 | ||
| Size ranges corresponding to recruits, prime, and old fish | Recruits <50 kg, prime 50–150 kg, old >150 kg | Obtained from discussions with ETBF industry members | |
| Decision-tree implementation | |||
| Number of years after which to invoke decision tree | 5 | ||
| Annual fishing mortality before invoking decision tree | 3 FMSY | High level of initial depletion to test the ability of the harvest strategy to rebuild the stock | |
| Frequency of RBC recalculation using decision tree | Annually | ||
| Sampling of catch | |||
| Perfect | 100% | ||
| Moderate, reflecting real-life sampling proportions | 70% | ||
| Poor | 2% | ||
| Population parameter | Base value | Reference | Definition and/or rationale |
|---|---|---|---|
| Steepness, h | 0.65 | Basson and Dowling (2007) | Ratio between recruitment (R) when spawning biomass is at 20% of unexploited spawning biomass (B0), and recruitment at the unexploited spawning biomass i.e. h = R(0.2 B0)/R(B0) |
| von Bertalanffy growth parameter, k | Male 0.1096, female 0.0815 | Young and Drake (2004) | Averaged so that there is no sexual dimorphism (in the reference case) |
| Natural mortality, M | 0.2 for all ages | ||
| Age at first maturity, tm | 9 | Young and Drake (2004) | |
| Standard deviation of the logarithm of the variability around the stock–recruitment curve, σR | 0.6 | Log(variation in year-class strength) | |
| Initial recruitment, InitR | 200 000 fish | ||
| Fishery parameter | |||
| Catchability, q | 1 × 10−5 | Yield magnitudes comparable with those historically reported in units of numbers per 1000 hooks | |
| Standard deviation of the logarithm of the variability in catch rate, σC | 0.2 | ||
| Size ranges corresponding to recruits, prime, and old fish | Recruits <50 kg, prime 50–150 kg, old >150 kg | Obtained from discussions with ETBF industry members | |
| Decision-tree implementation | |||
| Number of years after which to invoke decision tree | 5 | ||
| Annual fishing mortality before invoking decision tree | 3 FMSY | High level of initial depletion to test the ability of the harvest strategy to rebuild the stock | |
| Frequency of RBC recalculation using decision tree | Annually | ||
| Sampling of catch | |||
| Perfect | 100% | ||
| Moderate, reflecting real-life sampling proportions | 70% | ||
| Poor | 2% | ||
Size-specific catch rates (cpueprime, cpueold, cpuerecuits) were obtained by multiplying cpue by the proportion of fish in the respective weight categories defining recruits (<50 kg), prime (50–150 kg), and old fish (>150 kg), where weight is inferred from length using a power relationship. A sampling level of 70% of the total catch is assumed. Bias-free numbers at age in the samples were generated by sampling ages randomly without replacement from a vector of ages in the same proportions as those in the true catch at age. A multinomial random variable generated ages to which each fish in the sample was assigned. A size distribution was sampled from the catch-at-age vector by assuming that length at (quarterly) age for each gender follows a normal distribution.
In the reference scenario simulations (Table 2), the decision tree was invoked after 5 years of fishing at 3 FMSY. This high level of initial depletion was used to assess the ability of the decision-tree framework to recover a heavily depleted stock. Once implemented, the decision tree calculates the RBC in the first quarter of each year over the next 25 years.
Scenarios
Parameter values and the nature of the sensitivity tests undertaken were informed by a regional swordfish stock assessment (Kolody et al., 2006). Each scenario was run for 50 realizations, each with a set of random recruitment errors and random errors on catch rate (Table 2). Table 3 describes the performance indicators used to evaluate the fishery outcomes. Relative values are presented for all indicators; absolute values are irrelevant in this context, because the model used is not conditioned on data.
Summary of performance indicators used in the harvest strategy evaluation.
| Performance indicator | Abbreviation (axis headings) |
|---|---|
| Median spawning biomass (percentage of initial) over the period of interest | MedianSpBio |
| Lowest spawning biomass (percentage of initial) over the period of interest | LowSpBio |
| Final spawning biomass (percentage of initial) for the period of interest | FinalSpBio |
| Number of years spawning biomass drops <0.5 BMSY (a limit reference point) during the period of interest | BiodropLim |
| Number of years spawning biomass drops <1.2 BMSY (target reference point; BMEY proxy) during the period of interest | BiodropTarget |
| Median proportion of old fish over the period of interest | MedianPropOld |
| Final proportion of old fish for the period of interest | FinalPropOld |
| Median annual catch (as a percentage of MSY) over the period of interest | MedianCatch |
| Median annual cpue [as a percentage of the initial (virgin) value] over the period of interest | MedianCPUEnum |
| Average interannual variability in fishing mortality over the period of interest (effort is not an explicit quantity in these simulations) | relchangeF |
| Average interannual variability in catch over the period of interest | relchangeC |
| Performance indicator | Abbreviation (axis headings) |
|---|---|
| Median spawning biomass (percentage of initial) over the period of interest | MedianSpBio |
| Lowest spawning biomass (percentage of initial) over the period of interest | LowSpBio |
| Final spawning biomass (percentage of initial) for the period of interest | FinalSpBio |
| Number of years spawning biomass drops <0.5 BMSY (a limit reference point) during the period of interest | BiodropLim |
| Number of years spawning biomass drops <1.2 BMSY (target reference point; BMEY proxy) during the period of interest | BiodropTarget |
| Median proportion of old fish over the period of interest | MedianPropOld |
| Final proportion of old fish for the period of interest | FinalPropOld |
| Median annual catch (as a percentage of MSY) over the period of interest | MedianCatch |
| Median annual cpue [as a percentage of the initial (virgin) value] over the period of interest | MedianCPUEnum |
| Average interannual variability in fishing mortality over the period of interest (effort is not an explicit quantity in these simulations) | relchangeF |
| Average interannual variability in catch over the period of interest | relchangeC |
Summary of performance indicators used in the harvest strategy evaluation.
| Performance indicator | Abbreviation (axis headings) |
|---|---|
| Median spawning biomass (percentage of initial) over the period of interest | MedianSpBio |
| Lowest spawning biomass (percentage of initial) over the period of interest | LowSpBio |
| Final spawning biomass (percentage of initial) for the period of interest | FinalSpBio |
| Number of years spawning biomass drops <0.5 BMSY (a limit reference point) during the period of interest | BiodropLim |
| Number of years spawning biomass drops <1.2 BMSY (target reference point; BMEY proxy) during the period of interest | BiodropTarget |
| Median proportion of old fish over the period of interest | MedianPropOld |
| Final proportion of old fish for the period of interest | FinalPropOld |
| Median annual catch (as a percentage of MSY) over the period of interest | MedianCatch |
| Median annual cpue [as a percentage of the initial (virgin) value] over the period of interest | MedianCPUEnum |
| Average interannual variability in fishing mortality over the period of interest (effort is not an explicit quantity in these simulations) | relchangeF |
| Average interannual variability in catch over the period of interest | relchangeC |
| Performance indicator | Abbreviation (axis headings) |
|---|---|
| Median spawning biomass (percentage of initial) over the period of interest | MedianSpBio |
| Lowest spawning biomass (percentage of initial) over the period of interest | LowSpBio |
| Final spawning biomass (percentage of initial) for the period of interest | FinalSpBio |
| Number of years spawning biomass drops <0.5 BMSY (a limit reference point) during the period of interest | BiodropLim |
| Number of years spawning biomass drops <1.2 BMSY (target reference point; BMEY proxy) during the period of interest | BiodropTarget |
| Median proportion of old fish over the period of interest | MedianPropOld |
| Final proportion of old fish for the period of interest | FinalPropOld |
| Median annual catch (as a percentage of MSY) over the period of interest | MedianCatch |
| Median annual cpue [as a percentage of the initial (virgin) value] over the period of interest | MedianCPUEnum |
| Average interannual variability in fishing mortality over the period of interest (effort is not an explicit quantity in these simulations) | relchangeF |
| Average interannual variability in catch over the period of interest | relchangeC |
Simulations are not conditioned on historical catch data or the perceived status of the stock. Nor are the scenarios tuned to begin the application of the harvest strategy at a uniform level of stock depletion to facilitate a direct comparison of catch and biomass levels. Differing scenarios had various rates of recruitment, productivity, or initial fishing mortality, so the decision tree is initially implemented on varying levels of fishable biomass in different scenarios. This prevents a direct comparison of scenarios, but does not compromise evaluation of the performance of the decision tree, assessment of its ability to recover a heavily depleted stock, or determining the direction of change in catch and biomass under different conditions.
Reference case harvest strategy performance system dynamics
The following reference set of conditions, with variations on the decision tree, was used.
Full decision tree. This is the reference case harvest strategy.
No decision rule applied; stock fished at 3 FMSY for 30 years. This equates to an ongoing high level of fishing pressure with no management intervention.
Simple slope-to-target RBC rule, curtailing the decision tree after Level 1.
Decision-tree components related to cpuerecruits omitted from the decision tree, i.e. where applicable, cpuerecruits is assumed to be high and not decreasing.
Sensitivity tests
The 17 sensitivity tests presented are detailed in Table 4. They included examining the effect of increases in catchability, q, crossed with the assumed selectivity function, S, and the assumed natural mortality, M [Tests (viii)–(xv); Table 4], informed by the uncertainty described by the 2006 regional stock assessment (Kolody et al., 2006).
Description of key sensitivity tests and their rationale.
| Test | Description | Rationale |
|---|---|---|
| Reference | Full decision tree with reference values | |
| (i) | Standard deviation of variability around stock–recruitment curve, σR = 0.2 | Examine response to reduced uncertainty about stock–recruitment relationship |
| (ii) | Increasing the catchability: cumulative positive random increments of mean 4% every 5 years (standard deviation of variability = 0.1) together with the lognormal observation error on cpue (standard deviation of variability = 0.1) | Examine effect of catchability with random error; values based on notions of historical rates of increase in catchability |
| (iii) | Initial recruitment, InitR, halved to 100 000 fish during the first 5 years | Confers additional stress on population; can decision tree still recover the population? |
| (iv) | 2-year time-lag on cpue and size information | Realistic information flow rate given current time-lags in data processing and advance notice requirements on quota setting |
| (v) | Decision tree applied and RBC recalculated every 5 years | Confers economic stability to fishery by not changing RBC annually |
| (vi) | Alternative size ranges used in the decision tree: recruits <30 kg, prime 70–120 kg, old >170 kg | Narrower size ranges give a smaller catch sample. Non-continuous size classes; not all size information is included in the decision rule |
| (vii) | Spawning potential ratio (SPR) reference point calculations in the decision tree based on biological inputs that differ from the simulation system dynamics: M = 0.15; tm = 7 | Values used in the decision-tree calculations assume a more robust stock (lower natural mortality rate, younger age-at-maturity) than that being simulated. Does the decision tree respond as strongly given overly optimistic population dynamics? |
| (viii)–(xv) | No increase in catchability or 2% annual increase in catchability AND dome-shaped selectivity or increasing selectivity with age AND constant M-at-age or M-at-age vector | Examines combined effect of three main uncertainties: the level of catchability (halved here compared with scenario 2, given the strong effect of 4% annual alone), the nature of the selectivity curve and the age-specific natural mortality |
| Selectivity curves and natural mortality vectors taken from regional stock assessments (Kolody et al., 2006) | ||
| (xvi)–(xvii) | Steepness = 0.5; 0.9 | Examine sensitivity to assumed steepness in stock–recruitment relationship |
| Test | Description | Rationale |
|---|---|---|
| Reference | Full decision tree with reference values | |
| (i) | Standard deviation of variability around stock–recruitment curve, σR = 0.2 | Examine response to reduced uncertainty about stock–recruitment relationship |
| (ii) | Increasing the catchability: cumulative positive random increments of mean 4% every 5 years (standard deviation of variability = 0.1) together with the lognormal observation error on cpue (standard deviation of variability = 0.1) | Examine effect of catchability with random error; values based on notions of historical rates of increase in catchability |
| (iii) | Initial recruitment, InitR, halved to 100 000 fish during the first 5 years | Confers additional stress on population; can decision tree still recover the population? |
| (iv) | 2-year time-lag on cpue and size information | Realistic information flow rate given current time-lags in data processing and advance notice requirements on quota setting |
| (v) | Decision tree applied and RBC recalculated every 5 years | Confers economic stability to fishery by not changing RBC annually |
| (vi) | Alternative size ranges used in the decision tree: recruits <30 kg, prime 70–120 kg, old >170 kg | Narrower size ranges give a smaller catch sample. Non-continuous size classes; not all size information is included in the decision rule |
| (vii) | Spawning potential ratio (SPR) reference point calculations in the decision tree based on biological inputs that differ from the simulation system dynamics: M = 0.15; tm = 7 | Values used in the decision-tree calculations assume a more robust stock (lower natural mortality rate, younger age-at-maturity) than that being simulated. Does the decision tree respond as strongly given overly optimistic population dynamics? |
| (viii)–(xv) | No increase in catchability or 2% annual increase in catchability AND dome-shaped selectivity or increasing selectivity with age AND constant M-at-age or M-at-age vector | Examines combined effect of three main uncertainties: the level of catchability (halved here compared with scenario 2, given the strong effect of 4% annual alone), the nature of the selectivity curve and the age-specific natural mortality |
| Selectivity curves and natural mortality vectors taken from regional stock assessments (Kolody et al., 2006) | ||
| (xvi)–(xvii) | Steepness = 0.5; 0.9 | Examine sensitivity to assumed steepness in stock–recruitment relationship |
Description of key sensitivity tests and their rationale.
| Test | Description | Rationale |
|---|---|---|
| Reference | Full decision tree with reference values | |
| (i) | Standard deviation of variability around stock–recruitment curve, σR = 0.2 | Examine response to reduced uncertainty about stock–recruitment relationship |
| (ii) | Increasing the catchability: cumulative positive random increments of mean 4% every 5 years (standard deviation of variability = 0.1) together with the lognormal observation error on cpue (standard deviation of variability = 0.1) | Examine effect of catchability with random error; values based on notions of historical rates of increase in catchability |
| (iii) | Initial recruitment, InitR, halved to 100 000 fish during the first 5 years | Confers additional stress on population; can decision tree still recover the population? |
| (iv) | 2-year time-lag on cpue and size information | Realistic information flow rate given current time-lags in data processing and advance notice requirements on quota setting |
| (v) | Decision tree applied and RBC recalculated every 5 years | Confers economic stability to fishery by not changing RBC annually |
| (vi) | Alternative size ranges used in the decision tree: recruits <30 kg, prime 70–120 kg, old >170 kg | Narrower size ranges give a smaller catch sample. Non-continuous size classes; not all size information is included in the decision rule |
| (vii) | Spawning potential ratio (SPR) reference point calculations in the decision tree based on biological inputs that differ from the simulation system dynamics: M = 0.15; tm = 7 | Values used in the decision-tree calculations assume a more robust stock (lower natural mortality rate, younger age-at-maturity) than that being simulated. Does the decision tree respond as strongly given overly optimistic population dynamics? |
| (viii)–(xv) | No increase in catchability or 2% annual increase in catchability AND dome-shaped selectivity or increasing selectivity with age AND constant M-at-age or M-at-age vector | Examines combined effect of three main uncertainties: the level of catchability (halved here compared with scenario 2, given the strong effect of 4% annual alone), the nature of the selectivity curve and the age-specific natural mortality |
| Selectivity curves and natural mortality vectors taken from regional stock assessments (Kolody et al., 2006) | ||
| (xvi)–(xvii) | Steepness = 0.5; 0.9 | Examine sensitivity to assumed steepness in stock–recruitment relationship |
| Test | Description | Rationale |
|---|---|---|
| Reference | Full decision tree with reference values | |
| (i) | Standard deviation of variability around stock–recruitment curve, σR = 0.2 | Examine response to reduced uncertainty about stock–recruitment relationship |
| (ii) | Increasing the catchability: cumulative positive random increments of mean 4% every 5 years (standard deviation of variability = 0.1) together with the lognormal observation error on cpue (standard deviation of variability = 0.1) | Examine effect of catchability with random error; values based on notions of historical rates of increase in catchability |
| (iii) | Initial recruitment, InitR, halved to 100 000 fish during the first 5 years | Confers additional stress on population; can decision tree still recover the population? |
| (iv) | 2-year time-lag on cpue and size information | Realistic information flow rate given current time-lags in data processing and advance notice requirements on quota setting |
| (v) | Decision tree applied and RBC recalculated every 5 years | Confers economic stability to fishery by not changing RBC annually |
| (vi) | Alternative size ranges used in the decision tree: recruits <30 kg, prime 70–120 kg, old >170 kg | Narrower size ranges give a smaller catch sample. Non-continuous size classes; not all size information is included in the decision rule |
| (vii) | Spawning potential ratio (SPR) reference point calculations in the decision tree based on biological inputs that differ from the simulation system dynamics: M = 0.15; tm = 7 | Values used in the decision-tree calculations assume a more robust stock (lower natural mortality rate, younger age-at-maturity) than that being simulated. Does the decision tree respond as strongly given overly optimistic population dynamics? |
| (viii)–(xv) | No increase in catchability or 2% annual increase in catchability AND dome-shaped selectivity or increasing selectivity with age AND constant M-at-age or M-at-age vector | Examines combined effect of three main uncertainties: the level of catchability (halved here compared with scenario 2, given the strong effect of 4% annual alone), the nature of the selectivity curve and the age-specific natural mortality |
| Selectivity curves and natural mortality vectors taken from regional stock assessments (Kolody et al., 2006) | ||
| (xvi)–(xvii) | Steepness = 0.5; 0.9 | Examine sensitivity to assumed steepness in stock–recruitment relationship |
Results
Long-term behaviour
Projections were undertaken on a practical time-scale of 30 years. Long-term (100-year) realizations for the reference scenario were used to determine whether and at what level the population stabilizes (Figure 3). Under the decision tree, the population recovered within 5 years of implementation and overshot the spawning biomass corresponding to MSY or SPR40 before stabilizing at ∼20% above the target by year 30 (Figure 3a). Cpue- and size-based indicators showed a similar pattern of stabilizing well above the target, and the average catch continued to increase for the first 25 years after implementation. Over 100 years, the population stabilized after ∼70 years at levels above the target of 60% of the initial cpueprime, and size-based cpue levels at SPR40, the proxies for 0.4 B0 (Figure 3b). Meanwhile, catch varied below its target. This overly conservative performance of the decision tree was a result of the error imposed on cpue, which decouples its direct relationship with abundance. A single realization of an alternative reference simulation with no error on cpue and a smaller random error on recruitment (σR = 0.2) shows the population oscillating with decreasing amplitude before stabilizing at target levels (Figure 3c and d). This implies that the assessment logic applied is accurate and that error on cpue introduces a conservative bias. The oscillation observed, even when cpue is directly proportional to abundance, is to be expected: as the population recovers, the slope of recent cpueprime increases along with the size-based indicators. The RBC therefore increases until spawning biomass is affected adversely and begins to decline. However, there is a delay before this is reflected in the indicators, with the slope of the recent cpueprime becoming negative and size-based indicators dropping below their relevant threshold values. The catch pattern therefore lags that of spawning biomass and the size-based indicators. Over time, the magnitude of the oscillations decreases as the decision tree drives the population iteratively to its target equilibrium.
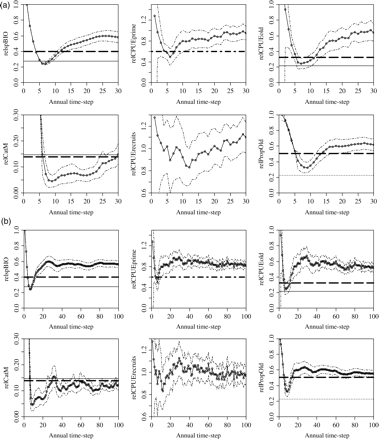
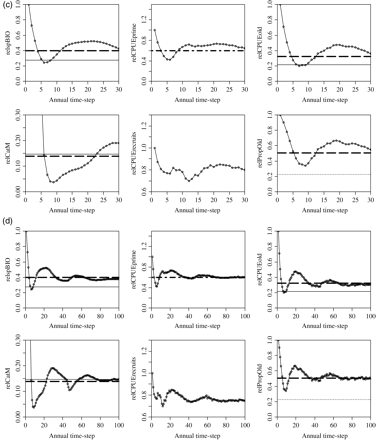
Time-series plots of mean and 95% confidence bounds for relative spawning biomass (axis label relspBIO), cpueprime (relCPUEprime), cpueold (relCPUEold), catch (relCatM), cpuerecruits (relCPUErecruits), and proportion of old fish (PropOld), over (a) 30, and (b) 100 years of simulation, for one realization of the reference scenario. The solid line is the average equilibrium level at MSY, the bold dashed line the level at SPR40, the fine dashed line the level at SPR20, and the bold, broken dashed (dot-dash) line the 60% initial cpueprime (slope-to-target rule target). These figures are also presented for a single realization of a scenario run without error imposed on cpue and with a lower (σR = 0.2) variability around the stock–recruitment curve in (c) and (d).
Decision-tree performance
Under continued fishing at 3 FMSY, the stock drops below 0.2 B0 after 30 years, but average catches over the last 20 years remain high (∼75% of MSY; Figure 4, scenario 2; Figure 5, triangle). In the absence of the size-based indicator rules used in the lower tiers of the decision tree, the slope-to-target equations (Figure 4, scenario 3; Figure 5, plus symbol) result in a higher spawning biomass (almost 0.4 B0) and a slightly lower final catch (∼70% of MSY) than when no decision rule is applied. The slope-to-target rule alone allows the stock to rebuild closer to a target level (0.6 × initial cpueprime; Figure 5). It reduces the catch strongly in the first 5 years (Figure 4a, MedianCatch), resulting in increased interannual variability in catch and fishing mortality, but over the final 20 years achieves comparable levels of catch with those under no decision rule (Figure 4b). Correspondingly, the proportion of old fish and the cpue are higher over years 10–30, and the spawning biomass is maintained above 50% BMSY in all years across all realizations (Figure 4b).

Boxplots for (a) the first 5 years after the decision tree was invoked (years 5–10), and (b) the last 20 years after the decision tree invoked (years 10–30), with the decision tree invoked in year 5, and scenarios 1–4.
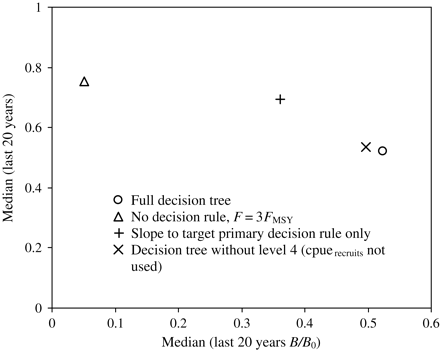
Median catch plotted against median spawning biomass, over years 10–30 of simulation, taken over 50 realizations, for scenarios 1–4.
When the full decision tree is applied, the highest spawning biomasses are achieved (>0.5 B0), though with reduced catches (∼50% of MSY). The population response is stronger in magnitude than that under the slope-to-target rule, with greater proportions of old fish and reduced incidences of the population dropping below its target biomass (Figure 4, scenario 1; Figure 5, open circle). Reductions in RBC are again greatest in the first 5 years after the implementation of the decision tree (Figure 4a, MedianCatch), suggesting that the decision tree responds conservatively when confronted with error on the cpue abundance index. It effectively recovers a heavily fished stock, but to above a target level, and at the expense of catch, especially in the early years of implementation. When compared with the slope-to-target decision rule alone, the decision-tree framework trades off marginally lower (∼20%) longer-term catches for an ∼15% higher spawning biomass at the end of the 30-year scenario.
Eliminating the use of the cpuerecruits indicator in the decision tree had little effect on the catch, but resulted in a slightly (∼3%) higher spawner biomass (Figure 5), suggesting that this indicator may be superfluous. Further investigation is warranted to determine whether this level of the decision tree is justified under alternative conditions.
Sensitivity analyses
The decision tree performed predictably in response to perturbations, alternative implementations, and input uncertainties. Generally, behaviour relative to the reference scenario lay along a continuum whereby catch was inversely related to spawning biomass. None of the scenarios tested resulted in the population dropping below the limit reference point of 20% of B0. Decreasing the level of random error in recruitment [Test (i)] resulted in little change across the indicators, but slightly higher spawning biomass and bigger catch (Table 5), suggesting that the decision tree acts more conservatively when confronted with more variability in the stock. Introducing positive 5-yearly ∼4% bias on the annual random error on cpue [Test (ii)] surprisingly did not have a strong effect, although the final spawner biomass was reduced by 10% and the median catch increased by 13%, with the spawner biomass on average dropping below the target level in just 3 years. The low-magnitude response was possibly because of the gradual (5-yearly) manifestation of the increase in catchability (q), together with the fact that its magnitude was subject to random variability, with additional error on cpue. Tests (viii)–(xv), which examined a fixed 2% annual increase in catchability, showed a greater effect on catch and spawning biomass and on the extent to which the population dropped below the target reference point.
Results from key sensitivity tests, with quantities expressed as medians over years 10–30 of the simulation, taken over 50 realizations.
| Test | MedianSpBio | LowSpBio | FinalSpBio | BiodropLim | BiodropTarget | MedianPropOld | FinalPropOld | MedianCatch | MedianCPUEnum | relchangeF | relchangeC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reference | 0.52 | 0.36 | 0.56 | 0 | 1.0 | 0.08 | 0.09 | 0.52 | 0.72 | 0.07 | 0.23 |
| (i) | 0.51 | 0.37 | 0.55 | 0 | 1.0 | 0.08 | 0.08 | 0.54 | 0.73 | 0.07 | 0.23 |
| (ii) | 0.47 | 0.32 | 0.46 | 0 | 3.0 | 0.08 | 0.08 | 0.65 | 0.76 | 0.13 | 0.28 |
| (iii) | 0.54 | 0.29 | 0.64 | 0 | 4.0 | 0.08 | 0.09 | 0.20 | 0.71 | 0.04 | 0.22 |
| (iv) | 0.38 | 0.27 | 0.51 | 0 | 9.5 | 0.06 | 0.07 | 0.56 | 0.59 | 0.20 | 0.56 |
| (v) | 0.26 | 0.22 | 0.33 | 0 | 20.0 | 0.05 | 0.06 | 0.71 | 0.49 | 0.13 | 0.06 |
| (vi) | 0.49 | 0.31 | 0.63 | 0 | 3.0 | 0.06 | 0.07 | 0.46 | 0.72 | 0.04 | 0.13 |
| (vii) | 0.47 | 0.36 | 0.51 | 0 | 1.5 | 0.07 | 0.08 | 0.59 | 0.68 | 0.07 | 0.23 |
| (viii) | 0.51 | 0.41 | 0.56 | 0 | 0.0 | 0.01 | 0.01 | 0.63 | 0.85 | 0.07 | 0.20 |
| (ix) | 0.54 | 0.36 | 0.58 | 0 | 1.0 | 0.06 | 0.06 | 0.49 | 0.77 | 0.07 | 0.22 |
| (x) | 0.49 | 0.48 | 0.51 | 0 | 0.0 | 0.02 | 0.02 | 0.71 | 0.80 | 0.05 | 0.14 |
| (xi) | 0.41 | 0.36 | 0.49 | 0 | 0.0 | 0.13 | 0.14 | 0.53 | 0.66 | 0.06 | 0.18 |
| (xii) | 0.37 | 0.31 | 0.35 | 0 | 5.5 | 0.01 | 0.01 | 0.93 | 0.98 | 0.16 | 0.28 |
| (xiii) | 0.36 | 0.24 | 0.31 | 0 | 11.5 | 0.04 | 0.04 | 0.79 | 0.86 | 0.16 | 0.31 |
| (xiv) | 0.41 | 0.35 | 0.35 | 0 | 0.0 | 0.02 | 0.02 | 1.09 | 0.96 | 0.10 | 0.18 |
| (xv) | 0.29 | 0.23 | 0.28 | 0 | 18.0 | 0.11 | 0.11 | 0.95 | 0.74 | 0.12 | 0.21 |
| (xvi) | 0.48 | 0.38 | 0.54 | 0 | 7.5 | 0.09 | 0.10 | 0.50 | 0.64 | 0.07 | 0.22 |
| (xvii) | 0.56 | 0.31 | 0.55 | 0 | 0.0 | 0.06 | 0.06 | 0.46 | 0.84 | 0.10 | 0.27 |
| Test | MedianSpBio | LowSpBio | FinalSpBio | BiodropLim | BiodropTarget | MedianPropOld | FinalPropOld | MedianCatch | MedianCPUEnum | relchangeF | relchangeC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reference | 0.52 | 0.36 | 0.56 | 0 | 1.0 | 0.08 | 0.09 | 0.52 | 0.72 | 0.07 | 0.23 |
| (i) | 0.51 | 0.37 | 0.55 | 0 | 1.0 | 0.08 | 0.08 | 0.54 | 0.73 | 0.07 | 0.23 |
| (ii) | 0.47 | 0.32 | 0.46 | 0 | 3.0 | 0.08 | 0.08 | 0.65 | 0.76 | 0.13 | 0.28 |
| (iii) | 0.54 | 0.29 | 0.64 | 0 | 4.0 | 0.08 | 0.09 | 0.20 | 0.71 | 0.04 | 0.22 |
| (iv) | 0.38 | 0.27 | 0.51 | 0 | 9.5 | 0.06 | 0.07 | 0.56 | 0.59 | 0.20 | 0.56 |
| (v) | 0.26 | 0.22 | 0.33 | 0 | 20.0 | 0.05 | 0.06 | 0.71 | 0.49 | 0.13 | 0.06 |
| (vi) | 0.49 | 0.31 | 0.63 | 0 | 3.0 | 0.06 | 0.07 | 0.46 | 0.72 | 0.04 | 0.13 |
| (vii) | 0.47 | 0.36 | 0.51 | 0 | 1.5 | 0.07 | 0.08 | 0.59 | 0.68 | 0.07 | 0.23 |
| (viii) | 0.51 | 0.41 | 0.56 | 0 | 0.0 | 0.01 | 0.01 | 0.63 | 0.85 | 0.07 | 0.20 |
| (ix) | 0.54 | 0.36 | 0.58 | 0 | 1.0 | 0.06 | 0.06 | 0.49 | 0.77 | 0.07 | 0.22 |
| (x) | 0.49 | 0.48 | 0.51 | 0 | 0.0 | 0.02 | 0.02 | 0.71 | 0.80 | 0.05 | 0.14 |
| (xi) | 0.41 | 0.36 | 0.49 | 0 | 0.0 | 0.13 | 0.14 | 0.53 | 0.66 | 0.06 | 0.18 |
| (xii) | 0.37 | 0.31 | 0.35 | 0 | 5.5 | 0.01 | 0.01 | 0.93 | 0.98 | 0.16 | 0.28 |
| (xiii) | 0.36 | 0.24 | 0.31 | 0 | 11.5 | 0.04 | 0.04 | 0.79 | 0.86 | 0.16 | 0.31 |
| (xiv) | 0.41 | 0.35 | 0.35 | 0 | 0.0 | 0.02 | 0.02 | 1.09 | 0.96 | 0.10 | 0.18 |
| (xv) | 0.29 | 0.23 | 0.28 | 0 | 18.0 | 0.11 | 0.11 | 0.95 | 0.74 | 0.12 | 0.21 |
| (xvi) | 0.48 | 0.38 | 0.54 | 0 | 7.5 | 0.09 | 0.10 | 0.50 | 0.64 | 0.07 | 0.22 |
| (xvii) | 0.56 | 0.31 | 0.55 | 0 | 0.0 | 0.06 | 0.06 | 0.46 | 0.84 | 0.10 | 0.27 |
Results from key sensitivity tests, with quantities expressed as medians over years 10–30 of the simulation, taken over 50 realizations.
| Test | MedianSpBio | LowSpBio | FinalSpBio | BiodropLim | BiodropTarget | MedianPropOld | FinalPropOld | MedianCatch | MedianCPUEnum | relchangeF | relchangeC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reference | 0.52 | 0.36 | 0.56 | 0 | 1.0 | 0.08 | 0.09 | 0.52 | 0.72 | 0.07 | 0.23 |
| (i) | 0.51 | 0.37 | 0.55 | 0 | 1.0 | 0.08 | 0.08 | 0.54 | 0.73 | 0.07 | 0.23 |
| (ii) | 0.47 | 0.32 | 0.46 | 0 | 3.0 | 0.08 | 0.08 | 0.65 | 0.76 | 0.13 | 0.28 |
| (iii) | 0.54 | 0.29 | 0.64 | 0 | 4.0 | 0.08 | 0.09 | 0.20 | 0.71 | 0.04 | 0.22 |
| (iv) | 0.38 | 0.27 | 0.51 | 0 | 9.5 | 0.06 | 0.07 | 0.56 | 0.59 | 0.20 | 0.56 |
| (v) | 0.26 | 0.22 | 0.33 | 0 | 20.0 | 0.05 | 0.06 | 0.71 | 0.49 | 0.13 | 0.06 |
| (vi) | 0.49 | 0.31 | 0.63 | 0 | 3.0 | 0.06 | 0.07 | 0.46 | 0.72 | 0.04 | 0.13 |
| (vii) | 0.47 | 0.36 | 0.51 | 0 | 1.5 | 0.07 | 0.08 | 0.59 | 0.68 | 0.07 | 0.23 |
| (viii) | 0.51 | 0.41 | 0.56 | 0 | 0.0 | 0.01 | 0.01 | 0.63 | 0.85 | 0.07 | 0.20 |
| (ix) | 0.54 | 0.36 | 0.58 | 0 | 1.0 | 0.06 | 0.06 | 0.49 | 0.77 | 0.07 | 0.22 |
| (x) | 0.49 | 0.48 | 0.51 | 0 | 0.0 | 0.02 | 0.02 | 0.71 | 0.80 | 0.05 | 0.14 |
| (xi) | 0.41 | 0.36 | 0.49 | 0 | 0.0 | 0.13 | 0.14 | 0.53 | 0.66 | 0.06 | 0.18 |
| (xii) | 0.37 | 0.31 | 0.35 | 0 | 5.5 | 0.01 | 0.01 | 0.93 | 0.98 | 0.16 | 0.28 |
| (xiii) | 0.36 | 0.24 | 0.31 | 0 | 11.5 | 0.04 | 0.04 | 0.79 | 0.86 | 0.16 | 0.31 |
| (xiv) | 0.41 | 0.35 | 0.35 | 0 | 0.0 | 0.02 | 0.02 | 1.09 | 0.96 | 0.10 | 0.18 |
| (xv) | 0.29 | 0.23 | 0.28 | 0 | 18.0 | 0.11 | 0.11 | 0.95 | 0.74 | 0.12 | 0.21 |
| (xvi) | 0.48 | 0.38 | 0.54 | 0 | 7.5 | 0.09 | 0.10 | 0.50 | 0.64 | 0.07 | 0.22 |
| (xvii) | 0.56 | 0.31 | 0.55 | 0 | 0.0 | 0.06 | 0.06 | 0.46 | 0.84 | 0.10 | 0.27 |
| Test | MedianSpBio | LowSpBio | FinalSpBio | BiodropLim | BiodropTarget | MedianPropOld | FinalPropOld | MedianCatch | MedianCPUEnum | relchangeF | relchangeC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reference | 0.52 | 0.36 | 0.56 | 0 | 1.0 | 0.08 | 0.09 | 0.52 | 0.72 | 0.07 | 0.23 |
| (i) | 0.51 | 0.37 | 0.55 | 0 | 1.0 | 0.08 | 0.08 | 0.54 | 0.73 | 0.07 | 0.23 |
| (ii) | 0.47 | 0.32 | 0.46 | 0 | 3.0 | 0.08 | 0.08 | 0.65 | 0.76 | 0.13 | 0.28 |
| (iii) | 0.54 | 0.29 | 0.64 | 0 | 4.0 | 0.08 | 0.09 | 0.20 | 0.71 | 0.04 | 0.22 |
| (iv) | 0.38 | 0.27 | 0.51 | 0 | 9.5 | 0.06 | 0.07 | 0.56 | 0.59 | 0.20 | 0.56 |
| (v) | 0.26 | 0.22 | 0.33 | 0 | 20.0 | 0.05 | 0.06 | 0.71 | 0.49 | 0.13 | 0.06 |
| (vi) | 0.49 | 0.31 | 0.63 | 0 | 3.0 | 0.06 | 0.07 | 0.46 | 0.72 | 0.04 | 0.13 |
| (vii) | 0.47 | 0.36 | 0.51 | 0 | 1.5 | 0.07 | 0.08 | 0.59 | 0.68 | 0.07 | 0.23 |
| (viii) | 0.51 | 0.41 | 0.56 | 0 | 0.0 | 0.01 | 0.01 | 0.63 | 0.85 | 0.07 | 0.20 |
| (ix) | 0.54 | 0.36 | 0.58 | 0 | 1.0 | 0.06 | 0.06 | 0.49 | 0.77 | 0.07 | 0.22 |
| (x) | 0.49 | 0.48 | 0.51 | 0 | 0.0 | 0.02 | 0.02 | 0.71 | 0.80 | 0.05 | 0.14 |
| (xi) | 0.41 | 0.36 | 0.49 | 0 | 0.0 | 0.13 | 0.14 | 0.53 | 0.66 | 0.06 | 0.18 |
| (xii) | 0.37 | 0.31 | 0.35 | 0 | 5.5 | 0.01 | 0.01 | 0.93 | 0.98 | 0.16 | 0.28 |
| (xiii) | 0.36 | 0.24 | 0.31 | 0 | 11.5 | 0.04 | 0.04 | 0.79 | 0.86 | 0.16 | 0.31 |
| (xiv) | 0.41 | 0.35 | 0.35 | 0 | 0.0 | 0.02 | 0.02 | 1.09 | 0.96 | 0.10 | 0.18 |
| (xv) | 0.29 | 0.23 | 0.28 | 0 | 18.0 | 0.11 | 0.11 | 0.95 | 0.74 | 0.12 | 0.21 |
| (xvi) | 0.48 | 0.38 | 0.54 | 0 | 7.5 | 0.09 | 0.10 | 0.50 | 0.64 | 0.07 | 0.22 |
| (xvii) | 0.56 | 0.31 | 0.55 | 0 | 0.0 | 0.06 | 0.06 | 0.46 | 0.84 | 0.10 | 0.27 |
Halving recruitment during the first 5 years [Test (iii)] resulted in a strong decrease in RBC, together with a slightly higher spawning biomass over the final 20 years (Table 5). Because of the delay before the fewer recruits enter the fishery and impact the decision-making process, reductions in catch and increases in biomass were greater only over the final 20 years. A 2-year time-lag on obtaining cpue and size information [Test (iv)] resulted in bigger catches and lower spawning biomass, and a greater incidence of the biomass dropping below the target reference point, because of the delayed response of the decision tree in detecting stock decline. Applying the decision tree every 5 years as opposed to annually [Test (v)] increased the catch by almost 20% and reduced interannual variability in the catch, and reduced the catch rate, median and final spawner biomass, and the proportion of old fish in the catch. As the decision tree was unable to react immediately to stock decline, constant catch quotas in the interim were high, particularly in the early years.
Restricting the size ranges used to define small, prime and old fish [Test (vi)] resulted in smaller catches and a slightly higher final spawner biomass, though a lower median spawner biomass, as a consequence of lower estimates of the proportion of old fish producing more conservative outcomes from the decision tree. Misspecification of natural mortality and age-at-maturity when calculating SPR [Test (vii)] resulted in a slightly lower spawning biomass but catches that were ∼7% higher (Table 5). As the cpueprime target is set at 60% of its initial level, the SPR value affects only the size indicator thresholds in the decision tree, and here their low values reflect the incorrect assumption of a more productive stock. Consequently, the decision tree behaves similarly to the scenario where only the primary decision rule was used, although spawning biomass was not reduced to the same extent. This suggests that size-based indicator thresholds still act to reduce the catch, though less effectively, and the stock still stabilizes, but at a lower level.
Tests (viii)–(xv) illustrate how factors in combination have the greatest effect on the population and the performance of the decision tree. An annual 2% increase in catchability [Tests (xii)–(xv); Table 5] consistently resulted in much higher catch but lower spawning biomass via indicators overestimating the condition of the stock, such that the population dropped, on average, below its target biomass in 18 years for Test (xv) (Table 5). A dome-shaped selectivity function [Tests (viii), (x), (xii), and (xiv)] generally yielded higher spawning biomasses and lower catches, by protecting larger fish so that the population did not drop below the target biomass for Test (xiv), but dropped below it on average for 18 years in Test (xv) without dome-shaped selectivity. When natural mortality increased with age [Tests (x), (xi), (xiv), and (xv)], spawning biomass was generally lower and catches higher. However, there were inconsistencies and exceptions in the trade-offs between catch and biomass when considering the effect of individual factors in combination with others, which underscores the importance of considering the interaction between factors when evaluating the direction of the response of the decision tree. When testing the decision rule in a model conditioned on data pertaining to the current stock status, it will be important to embrace the full range of uncertainties in combination when considering its performance.
Finally, the model was reasonably robust to changes in steepness. When steepness was reduced from 0.65 to 0.5 [Test (xvi)], spawner biomass was slightly reduced and the biomass dropped below the target in >7 years on average, although the final spawner biomass was only 2% less and other indicators were relatively unaffected (Table 5). When steepness was set at 0.9 [Test (xvii)], the median spawner biomass was slightly higher and the final spawner biomass 1% lower, but the catch was reduced by 6%.
Discussion
Harvest strategy evaluation
This pilot evaluation of harvest strategy was conducted to test whether the proposed ETBF harvest strategy performed as designed across a range of alternative conditions. The analyses were undertaken in a simplified manner; sources of error and bias were simple and known, confounding factors were limited to a maximum of three in any scenario, and the stock modelled was heavily and rapidly exploited before the implementation of the harvest strategy. However, our initial evaluation suggested that the decision tree has potential to produce management corrections across a range of modelled stock perturbations, fisheries dynamics, and data biases. On this basis, Kolody et al. (2009) undertook a fully data-conditioned harvest strategy evaluation of the decision tree across all main target ETBF species, and the approach has subsequently been implemented in the fishery.
Here, the decision tree adjusted catches rapidly during the first few years of its application, before stabilizing at a level of biomass above the target reference point and a level of catch well below MSY. Relative to the performance of the simpler slope-to-target primary decision rule used alone, the use of the decision tree acted more conservatively to increase the magnitude of initial management corrections, trading off an ∼20% reduction in catch for an ∼15% improvement in spawning biomass by the end of the 30-year scenario. The overconservative response of the rule was due to the error imposed on the catch rates, decoupling their direct relationship with abundance, as scenarios conducted without error stabilized at target levels. This suggests that the decision-tree logic is sound and, where uncertainty can be quantified, this bias could possibly be addressed by lowering the target level such that the realized indicator values approach the true target.
Interacting combinations of errors and uncertainties had the greatest effect on perturbing decision-tree performance, for which reason the results need to be interpreted with caution. That stated, a gratifying aspect of the decision-tree performance under initial evaluation was its precautionary behaviour, with greater uncertainty increasing its conservative bias, favouring lower catches and higher spawning biomasses. Additionally, misspecification of the biological parameters used to estimate the SPR reference points did not appear to drive ongoing long-term stock declines, the decision tree still stabilized the stock, and this was likely to have been as a result of the cpueprime target being set at 60% of its initial value.
Similarly the decision tree became more precautionary at low sampling levels; it should be emphasized, however, that these simulations did not include process error in selectivity. Additionally, the baseline assumption that 70% of the catch is measured is unlikely to be realistic for many fisheries. Further, it is seldom that the sampling process is truly random; for example, selectivity may vary interannually. Sampling uncertainty needs to be captured correctly when applying a decision rule dependent on size data. In the ETBF, processing practices result in individual weights being available for >90% of the catch for most species, so the 70% sampling assumption was considered conservative for this fishery. However, under the “poor” simulated sampling regimes (2% of the catch; Table 2), zero values of proportionold became common, resulting in the decision tree selecting precautionary management actions relative to true stock status. An advantage of this approach, therefore, is the explicit incentive it offers for fishers to engage in simple data-collection programmes aimed at documenting size-based catch rates. When fishing communities understand how simple catch data can improve the reliability of the harvest strategy, they are more likely to support cost-effective data-collection programmes (Prince et al., 2008).
Empirical vs. model-based indicators
There is growing awareness that while the problem of overfishing intensifies (Mullon et al., 2005), many approaches to addressing overfishing require scientific knowledge, datasets, and model structures beyond the capability of most fisheries (Walters and Pearse, 1996; Cochrane, 1999). This underscores the need to re-evaluate reliance on data-intensive stock assessment models and to re-assess the utility of empirical procedures for assessing fish stocks, based on simple indicators collected directly from the catch of a fishery.
Whereas empirical approaches to assessing fisheries relative to selected reference points lack the theoretical rigour associated with quantitative model-based approaches (Hilborn, 2002), our results suggest that it may be a pragmatic alternative for the ECTBF. First, being based on empirical indicators derived from local catches, the approach should be robust to uncertainty and changing knowledge of the linkages with the broader WCPO stocks. The Australian fishery would respond to the actual fishing pressure experienced by local stocks, rather than to impacts that might be imputed to exist by an international model-based assessment dependent on the assumed rates of mixing between ocean basins. Second, using existing data, the harvest strategy framework is relatively cost-neutral to implement. Third, empirical approaches have the advantage of simplicity, because the decision rule inputs are readily available, calculated with minimal technical expertise, and may be more readily understood and accepted (Hilborn, 2002). Sophisticated assessment models require a high level of expertise and expense to maintain and update. The associated reference points and decision rules are also difficult to explain to fishers with their black-box perception of the models on which they are based. The lack of an easily comprehended linkage between data inputs and model-derived indicators can lead to a sense of distrust and suspicion from industry. Less precise, yet relatively robust, empirical approaches based on simple indicators may be more easily understood by fishers, and hence more likely to receive industry support. Having presented the ETBF approach to industry groups and managers and commenced the implementation process, this strength is being emphasized by industry (B. Jefferies, Australian Tuna Boat Operators Association, pers. comm.).
Conversely, the simplicity of empirical approaches may be considered a weakness. Standardized catch rates have long been used as proxy abundance indicators, but the relationship between cpue and abundance can be tenuous, and data are often lacking to account for the differences in gear type and targeting practices that influence cpue trends (Paloheimo and Dickie, 1964; Harley et al., 2001; Walters, 2003). These variables also influence the size of fish being caught. This issue is especially pertinent in multispecies fisheries, such as the ETBF, where vessels can switch target species depending on resource availability and market conditions. This creates difficulties in monitoring trends in catch rate and size composition. However, in the absence of expensive fishery-independent surveys, model-based approaches also rely on these same potentially biased indices, and research into abundance indices needs to be maintained. We suggest that complicated model-based assessments should still be undertaken for those fisheries in which it is feasible, but the periodicity should be reduced, with the emphasis shifted to the quantification of operating models from which simple robust harvest strategies can be developed.
Scale-less stock assessment
Assumptions about the spatial scale of fish stocks are also critical to the accuracy of assessment modelling and management success (Orensanz and Jamieson, 1998; Prince, 2003; Hilborn et al., 2005). Hilborn et al. (2005) suggest that mismatch between the spatial scale of the resource and the scale of assessment and management is one of the three main causes for unexpected fisheries failure. Indeed, the uncertain linkages between the stocks fished by the Australian fleet in the CTS, and the stocks fished internationally across the broader WCPO, were a major driver for pursuing an empirical approach for the ETBF. Although the spatial complexity created by the highly migratory behaviour of tuna and billfish species was a driver for this work, there is growing appreciation that many fisheries are based on complex meta-populations, consisting of largely self-recruiting small-scale populations (Orensanz and Jamieson, 1998; Prince, 2003), or localized populations of juveniles and adults variably linked by complex flows of larvae (San Martín et al., 2010). For small-scale species, as well as highly migratory ones, inappropriate assumptions about the scale and structure of stocks can undermine sophisticated approaches to assessment and management. This problem underscores the need for harvest policies that are robust to alternative hypothesized model structures.
Our evaluation assumed that the harvest strategy was applied uniformly to a single unit stock. Although it is not necessary to resolve stock structure issues when applying the empirical decision tree, the results of our analysis are predicated on the assumption that all sources of fishing mortality can be modulated according to the decision rules of the harvest strategy evaluated. Other scenarios, not reported here, tested variations to the unit-stock assumption and predictably showed that the extent to which stocks are local determines the extent to which localized management will be successful. If the CTS stocks are strongly linked with the broader WCPO, the long-term sustainability of these resources will depend more on the functionality of the RFMO and the compliance of its members than on the local implementation of this harvest strategy. However, the Commonwealth Harvest Strategy Policy framework demands that local harvest strategy approaches provide the Australian Government with high moral ground from which precautionary management can be advocated for the entire WCPO.
If extended by local adaptation across the entire range of a fishery, rather than a single component within a fishery, the empirical approach presented here has potential to provide a cheaper, scale-less approach to stock assessment, matching local catch levels to local levels of reproductive potential, at the same time reducing the need for expensive information pertaining to the scale and structure of stocks. The complexities and expense of population-based modelling imposed by adult movement and juvenile migration might be largely circumvented if the approach was applied across all regions of a resource. Maintaining all components of a stock as sources rather than sinks may also avoid the “Tragedy of the Larval Commons” described by San Martín et al. (2010). However, we recognize that there are certain forms of non-stationary dynamics (e.g. recruitment regime shifts, interannual migration variability) that will severely challenge this harvest strategy (and complicated population models), and we encourage further development under more diverse conditions.
As this approach is based on detecting the impact of fishing on the size composition of a stock, it is not applicable universally. It would have limited applicability to fisheries based on a single pulse of fishing on a cohort at the end of a life cycle, such as on Pacific salmon. In a gauntlet-style fishery (Prince, 2005) where juvenile or subadult cohorts are fished only briefly, the size structure of the catch could not be used in the manner described here, although this approach might be adapted around a survey of the adult population. Despite the acknowledged exceptions, our approach offers an advance towards simpler, more cost-effective forms of assessment that may be applicable to many of the world's fisheries.
Supplementary material
Acknowledgements
We thank the members of the ETBF Harvest Strategy Working Group and industry members of the ETBF for their valuable input and feedback, and Keith Sainsbury and Ray Hilborn for feedback and support. Ana Parma and an anonymous reviewer kindly provided comprehensive comments on an earlier version of the manuscript. This work was funded by the Australian Fisheries Management Authority and CSIRO Wealth from Oceans Flagship.
References
Author notes
Present address: Indian Ocean Tuna Commission, Seychelles.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}