





Abstract
Feedback and change are basic features of ecosystems, something global change has highlighted. Changes in the physical environment will see shifts in species ranges, community compositions, and ultimately the form and function of ecosystem and the human societies that exploit them. What these shifts will be depends on which of the competing (and potentially counteracting) mechanisms dominate through space and time. Moreover, changes are unlikely to be simple or linear; there will be winners, losers, and surprises. It also means that management will be complex and non-stationary, presenting management, scientific, and statistical challenges.Fulton, E. A. 2011. Interesting times: winners, losers, and system shifts under climate change around Australia. – ICES Journal of Marine Science, 68: 1329–1342.
Introduction
The effect of cumulative human action on the climate and biosphere of Earth has the potential of affecting significantly many critical marine ecosystem services, in particular biodiversity and food security, as highlighted by the work of Cheung et al. (2008, 2009). IPCC (2007) scenarios for fossil-fuel emissions suggest that over the next 60–100 years, ocean surface temperatures are likely to warm by 1.8–4°C, the sea level is predicted to rise by 0.2–1.4 m (Meehl et al., 2005; IPCC, 2007; Rahmstorf, 2007), storms are likely to intensify, and ocean acidification could result in declines in pH of 0.14–0.5 (Caldeira and Wickett, 2005; IPCC, 2007). In addition, the projected rise in global population to 9 billion by 2050 (UN, 2009) could plausibly see 50–77% of the worlds population concentrated along coastal fringes (Small et al., 2000; CCSR, 2006), putting even more degradative pressure on coastal habitats.
Together, these shifting pressures are likely to result in significant shifts in species distributions (Cheung et al., 2009) and altered foraging and reproductive success (Bograd et al., 2009; Munday et al., 2009). The reduction in pH could also result in mortality of calcifying organisms in the benthos and plankton (Fabry et al., 2008; Hall-Spencer et al., 2008).
Since the release of the fourth assessment report by the IPCC (2007), there has been an increasing awareness of the topic, not only in scientific circles [where the number of papers referencing the topic has increased by 224% (web of knowledge search 26 May 2010)], but also among conservation-orientated non-government organizations, government and management agencies (many of which have initiated climate adaptation programmes or departments), and the public [with Google searches on climate change growing by 177% (Google trends http://www.google.com/trends?q=climate+change)]. There is now a keen interest among resource management groups about what ecological, economic, and social implications are associated with shifts in ecosystem structure and function induced by global change.
Until recently, most models [e.g. integrated assessment models (IAMs)] used to explore these issues have either used simple mathematical relationships to deal with environmentally driven damage to the economy or have been one-way coupled (where the conditions from a physical model force an economic model, or vice versa, with no dynamic feedback between them). IAMs represent key features of human systems (such as demography, energy use, technology, emissions, land use, and the economy) and simplified representations of the climate, ecosystems, and associated economic impacts (i.e. damage functions). The majority of IAMs that have been used to link changes in the climate with economic outcomes have used relatively simple damage functions that assume loss in productivity increases in proportion to some power of temperature change (Stanton et al., 2008). Little or no consideration is given to other routes of damage (e.g. changed patterns of rainfall, ocean acidification, sea-level rise, and inundation) or to feedbacks that could occur through a multitude of ecological pathways. There has also been a terrestrial focus, meaning that appreciation of the effect of global change on marine systems is not as well advanced (Richardson and Poloczanska, 2008), though many research programmes on the topic have been initiated in the past few years [e.g. the Bering Ecosystem Study and Bering Sea Integrated Ecosystem Research Program (BEST-BSIERP); Hollowed et al., 2009].
Those studies that have been undertaken in the marine realm have often been based around bioclimate envelope models. This kind of model gives insight into potential distributional shifts of species based on predicted shifts in suitable (or preferred) physical and biological conditions that have been identified from current species distributions (Pearson and Dawson, 2003). Although these models did not initially include interspecies relationships, adaptation or evolution, habitat dependence, phenology, or dispersal mechanisms, they are becoming increasingly sophisticated and the most recent marine examples do include at least some of these processes (e.g. population and dispersal dynamics were included in the model of Cheung et al., 2009).
A new class of model being applied to the question of the implications of global change for marine ecosystems and the societies that exploit them is an end-to-end (or whole-of-system) models. These mechanistic models attempt to integrate abiotic, biotic, and anthropogenic processes across different scales, allowing for dynamic two-way coupling (or interactions) between ecosystem components from microbes to top predators and the human sectors that exploit the system (Travers et al., 2007; Fulton 2010). These models are beginning to be coupled with regionally downscaled ocean–atmosphere models (e.g. the models being developed for BEST-BSIERP), though typically circulation, temperature, or production regimes are taken from these models and used to drive the bottom end of an end-to-end model (Brown et al., 2009). The models have usually only considered a single driver or correlated drivers (e.g. thermally driven changes in the environment), but cumulative impacts are likely to be critical regarding breaching ecosystem thresholds. This paper discusses a very early attempt to consider how multiple drivers might simultaneously act to reshape Australian marine ecosystems.
Methods
Because no single end-to-end model covers the entire expanse of Australian waters, seven existing regional models (summarized in Table 1 and described below) were modified to allow for representation of changes in temperature, salinity, pH, and dissolved oxygen regimes (which in turn could drive shifts in species distributions), sea level, storm frequency and intensity, the entry or exit of species with shifting distributions, and the location and nature of major coastal industries and regulatory systems. Between them, the models cover nearly two-thirds of Australia's EEZ, across the southeast (SE) of Australia and down the tropical coasts of both northeast (NE) and northwest (NW) Australia (Figure 1).
List of the three model types (with seven regional-scale implementations) used in the study with brief comments on modifications made to allow for consideration of climate change effects.
| Model | Model type | Regions covered | References | Modifications |
|---|---|---|---|---|
| Atlantis | Deterministic three-dimensional end-to-end model with explicit human behaviour (fisheries) | SE of Australia* | EAF et al. (2007) | To expand the anthropogenic components, Atlantis-SE was coupled with coastal industries and biophysical impacts model (see main text and Supplementary material for details) |
| InVitro | Agent-based end-to-end model with explicit human behaviour and multiple coastal industries | Ningaloo-Exmouth region* | EAF et al. (2009) | To cover the region seamlessly, the InVitro models were coupled spatially; they were also coupled with a coastal biophysical impacts model |
| Northwest shelf Australia* | Gray et al. (2006) and Little et al. (2006) | |||
| Ecospace | Trophodynamic two-dimensional model with F-based human activities (typically fisheries) | Great Barrier Reef* | Gribble (2003) | Each Ecospace model was coupled with a nutrient-phytoplankton-zooplankton-detritus to capture better environmentally driven changes in production and to coastal industries and biophysical impacts models. In the NW, the two Ecospace models (for the Pilbara and Gascoyne) were coupled spatially |
| Eastern Bass Strait† | Bulman et al. (2006) | |||
| Northwest shelf Australia† | Bulman et al. (2006) | |||
| Ningaloo-Exmouth region† | EAF (unpublished data) |
| Model | Model type | Regions covered | References | Modifications |
|---|---|---|---|---|
| Atlantis | Deterministic three-dimensional end-to-end model with explicit human behaviour (fisheries) | SE of Australia* | EAF et al. (2007) | To expand the anthropogenic components, Atlantis-SE was coupled with coastal industries and biophysical impacts model (see main text and Supplementary material for details) |
| InVitro | Agent-based end-to-end model with explicit human behaviour and multiple coastal industries | Ningaloo-Exmouth region* | EAF et al. (2009) | To cover the region seamlessly, the InVitro models were coupled spatially; they were also coupled with a coastal biophysical impacts model |
| Northwest shelf Australia* | Gray et al. (2006) and Little et al. (2006) | |||
| Ecospace | Trophodynamic two-dimensional model with F-based human activities (typically fisheries) | Great Barrier Reef* | Gribble (2003) | Each Ecospace model was coupled with a nutrient-phytoplankton-zooplankton-detritus to capture better environmentally driven changes in production and to coastal industries and biophysical impacts models. In the NW, the two Ecospace models (for the Pilbara and Gascoyne) were coupled spatially |
| Eastern Bass Strait† | Bulman et al. (2006) | |||
| Northwest shelf Australia† | Bulman et al. (2006) | |||
| Ningaloo-Exmouth region† | EAF (unpublished data) |
Note that end-to-end means a model spanning physics, the foodweb, habitats, and human industries (especially fisheries).
*Models were the primary models run.
†Models were run for comparative purposes only.
List of the three model types (with seven regional-scale implementations) used in the study with brief comments on modifications made to allow for consideration of climate change effects.
| Model | Model type | Regions covered | References | Modifications |
|---|---|---|---|---|
| Atlantis | Deterministic three-dimensional end-to-end model with explicit human behaviour (fisheries) | SE of Australia* | EAF et al. (2007) | To expand the anthropogenic components, Atlantis-SE was coupled with coastal industries and biophysical impacts model (see main text and Supplementary material for details) |
| InVitro | Agent-based end-to-end model with explicit human behaviour and multiple coastal industries | Ningaloo-Exmouth region* | EAF et al. (2009) | To cover the region seamlessly, the InVitro models were coupled spatially; they were also coupled with a coastal biophysical impacts model |
| Northwest shelf Australia* | Gray et al. (2006) and Little et al. (2006) | |||
| Ecospace | Trophodynamic two-dimensional model with F-based human activities (typically fisheries) | Great Barrier Reef* | Gribble (2003) | Each Ecospace model was coupled with a nutrient-phytoplankton-zooplankton-detritus to capture better environmentally driven changes in production and to coastal industries and biophysical impacts models. In the NW, the two Ecospace models (for the Pilbara and Gascoyne) were coupled spatially |
| Eastern Bass Strait† | Bulman et al. (2006) | |||
| Northwest shelf Australia† | Bulman et al. (2006) | |||
| Ningaloo-Exmouth region† | EAF (unpublished data) |
| Model | Model type | Regions covered | References | Modifications |
|---|---|---|---|---|
| Atlantis | Deterministic three-dimensional end-to-end model with explicit human behaviour (fisheries) | SE of Australia* | EAF et al. (2007) | To expand the anthropogenic components, Atlantis-SE was coupled with coastal industries and biophysical impacts model (see main text and Supplementary material for details) |
| InVitro | Agent-based end-to-end model with explicit human behaviour and multiple coastal industries | Ningaloo-Exmouth region* | EAF et al. (2009) | To cover the region seamlessly, the InVitro models were coupled spatially; they were also coupled with a coastal biophysical impacts model |
| Northwest shelf Australia* | Gray et al. (2006) and Little et al. (2006) | |||
| Ecospace | Trophodynamic two-dimensional model with F-based human activities (typically fisheries) | Great Barrier Reef* | Gribble (2003) | Each Ecospace model was coupled with a nutrient-phytoplankton-zooplankton-detritus to capture better environmentally driven changes in production and to coastal industries and biophysical impacts models. In the NW, the two Ecospace models (for the Pilbara and Gascoyne) were coupled spatially |
| Eastern Bass Strait† | Bulman et al. (2006) | |||
| Northwest shelf Australia† | Bulman et al. (2006) | |||
| Ningaloo-Exmouth region† | EAF (unpublished data) |
Note that end-to-end means a model spanning physics, the foodweb, habitats, and human industries (especially fisheries).
*Models were the primary models run.
†Models were run for comparative purposes only.
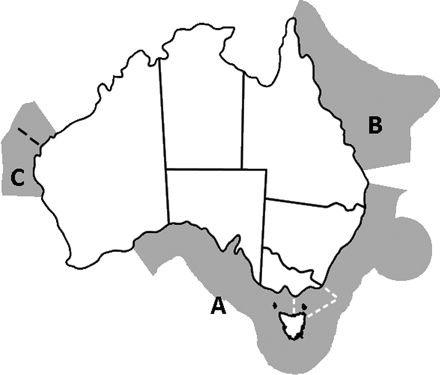
Areas modelled in the current study: (A) SE of Australia (white dashed line marks the boundary of the Eastern Bass Strait region), (B) Great Barrier Reef and (C) northwest of Australia (consisting of the Northwest shelf and Ningaloo-Exmouth regions—the black dashed line marks the boundary between these two regions).
Three modelling frameworks were used as a basis for the model systems: Atlantis (Fulton et al., 2005), InVitro (Gray et al., 2006), and Ecospace (Walters et al., 1999). All these frameworks have the potential of covering all the main biological components of marine ecosystems, from primary production to top predators. The biggest differences between the models are in how they represent the different components. Before discussing how the models were used, a brief overview is given of the main structure and assumptions of the models and the specific implementations used to represent the different Australian areas. Each model is the outcome of many years work in each region, and interested readers are encouraged to seek extra detail in the references provided.
Atlantis
The model for SE Australia is a modified form of Atlantis-SE (Fulton et al., 2007), which was originally developed as the basis for a whole-of-ecosystem management strategy evaluation in support of a strategic restructuring of SE Australian federal fisheries. The model has also been used to look at general fisheries and climate-related questions, such as multispecies maximum sustainable yield (MMSY; Worm et al., 2009) and the efficacy of ecological indicators of the effects of fishing (Branch et al., 2010). This model has the broadest spatial scale (covering 3.7 million km2) and quite complex ecology, including a size-resolved microbial web; nutrient, light, and space-based primary production; 37 age-structured ecological groups (from forage fish to top predators, listed in Supplementary Table A1), some resolved to the species level; multiple genetic stocks per group; and shifting climate-related environmental drivers of physiology and reproduction. There is also explicit representation of recreational and commercial fishing fleets, with the latter resolved to subfleets defined by homeports, crew, and vessel sizes; and driven by social and economic drivers that determine investment, disinvestment, quota trading, information updating, and effort allocation.
All Atlantis models are built upon a largely deterministic, (coarsely) spatially resolved three-dimensional biophysical submodel that tracks the nutrient (usually nitrogen and silica) flows through the main biological groups of the system of interest on a 6–24-h time-step (typically). The identity and configuration of these groups is defined based on a wide range of options, including some form of representation of consumption, production, waste cycling, movement, recruitment, and habitat dependence (example formulations are given in the Supplementary material). The physical environment (including water and substratum properties and processes) is represented using irregular polygons matched to the major geographical and bioregional features of the simulated marine system. The anthropogenic components of the model represent the impact of pollution and coastal development, but are focused on the detailed dynamics of fishing fleets and management regimes. The exact form used can again be drawn from a wide list of options, but for Atlantis-SE includes explicit socio-economically driven effort allocation (example formulations again given in the Supplementary material) and regulation based on gear restrictions, individual transferable quotas, spatial and temporal zoning, discarding restrictions, size limits, bycatch mitigation, and dynamic reference points and decision rules.
It is quite difficult to find sufficient data to fit fully such large models, but in this case, there has been extensive calibration ecologically (vs. 36, 20–90-year catch history time-series and sporadic scientific surveys) and anthropogenically (though in this case only 7 years of data were available for testing, after removing the first 10-year section of the available effort time-series as a training dataset). Using a simple implementation of pattern-orientated modelling (which simultaneously fits the entire model against data from multiple datasets; Kramer-Schadt et al., 2007), bounding parametrizations were found that produce equally plausible modelled systems given the available data and alternative possible system structures. These parametrizations were then all carried forward in the various simulations done for the final analyses.
InVitro
The NW region was represented by an InVitro model, formed by combining InVitro-NWS (Little et al., 2006; McDonald et al., 2006, 2008) and InVitro-Ningaloo (Fulton et al., 2009), which were initially developed for evaluating multiple use management strategies in the Pilbara and the Gascoyne regions of NW Australia. InVitro (Gray et al., 2006) is a hybrid agent-based modelling platform, where the behaviour and representation of each of the biological or anthropogenic submodels or agents is specific to their type (a classification of the agent types and some examples are given in Supplementary Table A1). For example, biophysically it couples classical dynamic differential equation and metapopulation models for lower trophic levels and habitats with decision-based agents for higher trophic levels, species of conservation concern (e.g. whales and whale sharks), and human sectors. Similarly, anthropogenic sectors are represented in a range of ways. Impacts of commercial industries, such as nutrient pollution, salt extraction, shipping, ports, coastal and urban development (including residential, service, industrial, and retail sectors), road transport systems (e.g. road trains), and the oil and gas sector are represented using differential equations or cellular automata (White et al., 2000) that mimic the impacts, activities, and management strategies of these industries. In contrast, coastal agriculture, fisheries, and tourism are much more detailed, using true individual-based models, with individuals or small groups explicitly making decisions about site-specific activities, based on simple economic and mechanical drivers; Kalman filters are used in these agent types to represent learning and updating of individual knowledge and experience (Gray et al., 2006). Monitoring and management associated with each of the sectors is also explicitly represented. As with Atlantis, a form of pattern-orientated modelling was used to produce bounding parameter sets that produce plausible model system states. A wide range of parametrizations was retained in this case, because of a lack of long-term time-series to constrain tightly possible parametrizations and the stochastic nature of the model, which made it harder to distinguish between alternative parametrizations with similar model skill.
Ecospace
The Great Barrier Reef (GBR) region was represented using a modified Ecospace model based on Gribble (2003). Ecospace is an explicitly spatial form of the Ecopath with Ecosim (EwE) modelling approach (Walters et al., 2000; Christensen and Walters, 2004), which uses biomass pools to represent the ecological components of the system. EwE can include age structure of key groups—using life-history stanzas (Walters et al., 2008)—and this was the case for two groups in this model (mangrove jacks and small schooling fish, updated to this form from an earlier age-structured method used in Gribble, 2003). To model the spatio-temporal dynamics of the system, Ecospace applies a set of differential equations (an example of the form is given in the Supplementary material) across a regular grid; with movement between cells based on a gradient model between sites of good and poor habitat or prey availability (though seasonal migration and particle-like individual-based movement options are also available; Walters et al., 2010). The non-spatial forms have been used widely and they have yielded good insights into system function and fisheries management issues (Robinson and Frid, 2003). Ecospace has not been as widely used, but given the spatial differential in potential impacts evident in the downscaled global climate model (GCM) products, explicit spatial handling were considered critical; in particular, because it allowed for consideration of spatial dispersal (and how this might change over time in response to environmentally driven changes in habitat and prey fields) and variations in productivity. This variability was captured via dynamic (two-way) coupling of ECOSPACE with a spatially resolved, environmentally driven, nutrient–phytoplankton–zooplankton–detritus (NPZD) model. Further details of this coupling are given below in the section on representing biophysical change.
The GBR Ecospace model of Gribble (2003) has 32 groups (listed in Supplementary Table A1), including two forms of detritus (with fisheries discards separate to other forms of detritus), pelagic and benthic primary producers, planktonic as well as benthic invertebrates (with penaeid prawns pulled out to a species level), 23 fish groups (some with age structure) and species of conservation concern, such as dugongs, large sea turtles, and seabirds. Line, trawl, and gillnet fisheries are explicitly represented in the model. The model was originally used to consider the implications of alternative management options for the prawn trawl fishery within the GBR, but has also been used to consider the implications of different levels of spatial zoning (Gribble, 2007, 2009) and more general issues, such as MMSY (Worm et al., 2009) and the efficacy of mean trophic level of the catch as a biodiversity indicator (Branch et al., 2010). This model was fitted to fisheries time-series and its spatio-temporal predictions have been validated against independent survey datasets (Gribble, 2009).
For comparative purposes, Ecospace models (modified in the same way as for the GBR; e.g. coupled with NPZD production models) for the SE and NW regions were run in parallel with the Atlantis and InVitro models (these are indicated with a dagger in Table 1). The EwE model for SE Australia is a shelf model of Eastern Bass Strait (a well-studied area that is key to the fisheries of the SE region and has biophysical features representative of those found across the broader region) first developed by Bulman et al. (2006). It contains 55 living groups and two forms of detritus (Supplementary Table A1), with a focus on the demersal fish community. The model has been fitted formally to 10-year time-series of data (for each of 42 groups) and has been used previously to consider a range of fisheries management issues, including ones specific to that area (Bulman et al., 2006), but also broader ones to do with fishing of forage fish, MMSY (Worm et al., 2009), and the usefulness of mean trophic level as a reliable measure of marine biodiversity (Branch et al., 2010).
The NW Ecospace model is formed from a model of the Pilbara shelf region of Australia (Bulman, 2007) and an unpublished model of the Ningaloo Reef and Exmouth Gulf region (Supplementary Table A1). Neither of these models has been fitted formally to continuous time-series (because none exists), but the spatio-temporal patterns they predict have been validated against data from scientific surveys collected sporadically in the region over the past 40 years. As for the SE model, these models have been used to look at management and development issues specific to the regions of the model, as well as more general fisheries concepts like MMSY and mean trophic level of the catch.
Ecosim (non-spatial) versions of the GBR, SE, and NW shelf models were also used in an earlier study that considered potential effects of climate driven changes in productivity (Brown et al., 2009).
Representing biophysical change
The Atlantis-SE currents (in the form of bulk exchanges) and water column properties were estimated using the Commonwealth Scientific and Industrial Research Organisation (CSIRO) 0.1° × 0.1° ocean forecasting model (OFAM) conditioned on the CSIRO Mark 3.5 coupled atmosphere–ocean global circulation model [a summary of these models is provided in Supplementary material; full details are available in Gordon et al. (2010) and Oke et al. (2005)]. The IPCC SRES A2 emission scenario (Nakicenovic and Swart, 2000) was used for the climate projections. The A2 scenario was chosen because it is consistent with the current levels of emissions (Rahmstorf et al., 2007), it is conservative regarding the level of global economic growth (Van Vuuren and O'Neill, 2006), and the broad-scale heterogeneity assumed in the scenario is most consistent with the heterogeneous dynamics of the anthropogenic components of Atlantis-SE and the coupled industries model. Storm events were represented via simple-impact models (where damage was applied to appropriate model components sitting under the footprint of the storm in proportion to the storm's intensity); and sea-level rise was also treated simply—via gradual changes in vertical layer depths and associated rates of sedimentation and inundation; mechanisms to allow for the shoreward retreat of coastal habitats were also incorporated. These processes were parametrized from the literature (Harmlien-Vivien and Laboute, 1986; Ellison and Stoddart, 1991; Dollar and Tribble, 1993; French, 1993; Michener et al., 1997; Short and Neckles, 1999; Nicholls, 2002, 2004; Reed, 2002; Scavia et al., 2002; McInnes et al., 2003; Williams et al., 2003; Crooks, 2004; Fourqurean and Rutten, 2004; Nicholls and Lowe, 2004; Zhang et al., 2004; Cruz-Palacios and van Tussenbroek, 2005; Feagin et al., 2005; Gardner et al., 2005; Church et al., 2006; Ericson et al., 2006; Harley et al., 2006) and conversations with regional geomorphology experts. All these changes (summarized in Table 2) then formed the basis of the abiotic environment that was acted on by the dynamic Atlantis components.
Summary of abiotic and global economic conditions used to set the broad context and to drive the end-to-end models (taken from IPCC SRES A2 scenario).
| Global system feature | Value in 2050 |
|---|---|
| Global properties | |
| Global GDP ($USa 1 000 billion year–1) | 59–111 |
| Global population (billion) | 9.7–11.3 |
| Regional heterogeneity | High |
| Economic development | Moderate |
| Global economic growth rate (%) | 1.0 |
| CO2 concentration (ppm) | 536 |
| Global temperature change (°C) | +1.2–1.8 |
| Global sea-level rise (m) | 0.1–0.25 |
| Global ocean pH | 7.9 |
| Australian properties | |
| Sea surface temperature change (°C) | +0.8–4.0 |
| pH change | –0.14 to –0.19 |
| Storm frequency and intensity | No increase in frequency, but average intensity increases |
| Precipitation | Up to 20% decrease in east–SE, potentially small increase in the north |
| Fires | More intense and frequent in SE |
| Australian population outside modelled coastal zone (million) | 3.5–4.8 |
| Global system feature | Value in 2050 |
|---|---|
| Global properties | |
| Global GDP ($USa 1 000 billion year–1) | 59–111 |
| Global population (billion) | 9.7–11.3 |
| Regional heterogeneity | High |
| Economic development | Moderate |
| Global economic growth rate (%) | 1.0 |
| CO2 concentration (ppm) | 536 |
| Global temperature change (°C) | +1.2–1.8 |
| Global sea-level rise (m) | 0.1–0.25 |
| Global ocean pH | 7.9 |
| Australian properties | |
| Sea surface temperature change (°C) | +0.8–4.0 |
| pH change | –0.14 to –0.19 |
| Storm frequency and intensity | No increase in frequency, but average intensity increases |
| Precipitation | Up to 20% decrease in east–SE, potentially small increase in the north |
| Fires | More intense and frequent in SE |
| Australian population outside modelled coastal zone (million) | 3.5–4.8 |
aThis is 1990 USD.
Summary of abiotic and global economic conditions used to set the broad context and to drive the end-to-end models (taken from IPCC SRES A2 scenario).
| Global system feature | Value in 2050 |
|---|---|
| Global properties | |
| Global GDP ($USa 1 000 billion year–1) | 59–111 |
| Global population (billion) | 9.7–11.3 |
| Regional heterogeneity | High |
| Economic development | Moderate |
| Global economic growth rate (%) | 1.0 |
| CO2 concentration (ppm) | 536 |
| Global temperature change (°C) | +1.2–1.8 |
| Global sea-level rise (m) | 0.1–0.25 |
| Global ocean pH | 7.9 |
| Australian properties | |
| Sea surface temperature change (°C) | +0.8–4.0 |
| pH change | –0.14 to –0.19 |
| Storm frequency and intensity | No increase in frequency, but average intensity increases |
| Precipitation | Up to 20% decrease in east–SE, potentially small increase in the north |
| Fires | More intense and frequent in SE |
| Australian population outside modelled coastal zone (million) | 3.5–4.8 |
| Global system feature | Value in 2050 |
|---|---|
| Global properties | |
| Global GDP ($USa 1 000 billion year–1) | 59–111 |
| Global population (billion) | 9.7–11.3 |
| Regional heterogeneity | High |
| Economic development | Moderate |
| Global economic growth rate (%) | 1.0 |
| CO2 concentration (ppm) | 536 |
| Global temperature change (°C) | +1.2–1.8 |
| Global sea-level rise (m) | 0.1–0.25 |
| Global ocean pH | 7.9 |
| Australian properties | |
| Sea surface temperature change (°C) | +0.8–4.0 |
| pH change | –0.14 to –0.19 |
| Storm frequency and intensity | No increase in frequency, but average intensity increases |
| Precipitation | Up to 20% decrease in east–SE, potentially small increase in the north |
| Fires | More intense and frequent in SE |
| Australian population outside modelled coastal zone (million) | 3.5–4.8 |
aThis is 1990 USD.
The same approach was taken to represent abiotic shifts, storms, and sea-level rise in the InVitro model. For the Ecospace models, simple NPZD models were created (using the Atlantis model framework) for the different regions. These NPZD models were driven by downscaled abiotic drivers in the same way as in the other models; they were also (two-way) coupled dynamically with the Ecospace models. The coupler used a simple interpolation from the Atlantis polygon map to the Ecospace grid. The coupler delivers biomass values from the NPZD to Ecospace, returning predation mortality rates on the plankton groups and detrital production by the groups within the Ecospace model. This transfer occurred during each Ecospace time-step (which is a month long), but for numerical stability the returns were evenly smeared over the next month's worth of NPZD time-steps. Although crude, this approach resulted in a production model similar in spatial patterns, realized biomass, and production levels to that of more sophisticated biogeochemical models (e.g. the coupled GCM-NPZ model described in Brown et al., 2009) and better captured shifts in the planktonic and primary producer groups than the standard Ecospace representations could, given the many changing abiotic factors that effected the lower trophic levels.
Representing change in the human systems
Fewer drivers were used to define the scenario of change from a social and economic perspective than for the abiotic factors. The social and economic changes seen in the simulations largely result from the dynamics of explicit anthropogenic submodels. However, the large contribution of exports to the Australian economy (and the large export markets that support many Australian industries) means that the general form of the global economy is needed as a model driver. The A2 scenario was chosen to provide this global context for two reasons. First, the economic projections are conservative compared with both the other scenarios and newer projections (Van Vuuren and O'Neill, 2006). Second, preliminary results indicated that realized rates of economic growth in the simulations were slower than assumed in the SRES A1 and B1 scenarios and that heterogeneity remained between sectors and regions (although this was only within the Australian model domains, it was assumed that this differential could also potentially result globally, if that were explicitly represented too). This is most consistent with the SRES A2 scenario. Consequently, the A2 emission scenario was chosen as the abiotic and broad economic context (summarized in Table 2) for this work.
Regarding dynamic model components for Atlantis-SE and InVitro, there were quite detailed sector and regulatory models. As discussed earlier, in InVitro, all the major coastal industries are represented explicitly and change in these industries in response to the biophysical changes is simply played out. Human responses also played out in Atlantis; however, the majority of the focus in Atlantis is on fisheries, with other sectors represented via an impact model. To make this impact model more responsive (i.e. more dynamic than a fixed constant or linearly changing level of impact), it was coupled with a dynamic stocks-and-flows representation of coastal sectors to determine the magnitude of potential impacts (primarily in the form of changed flows from estuaries, including the magnitude and seasonal patterns of the river flow, but also pollutant content and salinity levels; levels of outflows of dissolved nutrients from catchments or outfalls; and substratum available for coastal habitats). This simple economics-based representation of coast industries allows for shifting demography, which effects available labour sources and demands on services, innovation, and technological uptake (which changes costs, prices, and efficiencies) and shifts in infrastructure (capacity, costs, and maintenance)—readers unfamiliar with system dynamics approaches can find a useful introduction to the concepts in Morecroft (2007). The formulations used in this study draw heavily on the representation of coastal industries in InVitro (in turn based heavily on White et al., 2000; Waddell et al., 2003) and the representation of technological shifts and inter-industry structures captured in Pant (2007). In all these representations, a temperature-dependent damage function is used to link the biophysical and human spheres. Further feedbacks exist in the models via fish production, coastal habitats, nutrient cycling, the status of charismatic species, and general habitability.
In contrast to the other models, which contain detailed social and economic submodels, the Ecospace models used simpler fleet dynamics models and relatively static forms of management. Given the high degree of uncertainty associated with all these models (Atlantis, InVitro, and Ecospace), alternative parametrizations were used to capture a range of levels of vulnerability and adaptive capacity.
Simulations and indicators
The models were run from a state representing the period 2010–2060. This end date is arbitrary and it reflects the fact that beyond this date the Atlantis-SE model became numerically unstable for reasons explained further below. During this projection period, all the ecological and socio-economic components were able to react dynamically to their circumstances—within the constraints of the model formulations.
Given the uncertainty in the model predictions and the fact that the models do not include dynamic (shifting value) parametrizations (or other means of representing evolution of shifts in diversity within functional groups), aggregate results rather than results per group were considered the most reliable indicators of potential shifts within the ecosystems (my previous experience indicates that responses of broad types of organism are far less uncertain regarding general trends and gross magnitude of any change than specifics for individual groups). Consequently, only changes in the relative biomass of primary producers, detritus, plankton, other pelagic invertebrates (e.g. jellyfish and squid), benthic invertebrates (both sessile and mobile), pelagic fish, demersal fish, and top predators are reported. Similarly, for socio-economic components, the results have been aggregated to give the relative change in the value of small-scale fisheries (for both small owner operators and recreational or artisanal fisheries) and large-scale commercial fisheries (particularly operations using large boats where the captains are not company owners).
Consideration of MMSY
As a complement to the predictions made about the future trajectories of the systems, the models were also used to calculate what MMSY would look like in the end-state ecosystems. The approach used was the same as documented for Worm et al. (2009), but instead of using the model systems with a 2000–2010 climate state as the basis for the analysis, the 2050–2060 climate state was used instead.
Results
Predicted biophysical changes
For NE and NW Australia, the models indicated increases of 5–22% in the relative biomass of plankton groups, 20–105% increases for other pelagic invertebrates, 21–880% for pelagic fish biomass, and a decline in demersal (often habitat-associated) fish of 8–35% (Table 3). The exact magnitude of these changes differed between systems, but the general patterns were similar. Although specifics varied between systems, the general reasons for these results are consistent—pelagic groups appeared to benefit more often than they suffered from changed water column properties, they were also able to move more freely to avoid any poor conditions; in addition, their life histories often allowed for more rapid responses and they seemed to have greater buffer capacity in their diets, switching composition more rapidly than is seen among the demersal groups. Furthermore, the demersal groups had more diet components degraded by changed environmental conditions; they therefore had less scope for focusing on resilient prey groups. The habitat association of many of the demersal groups also constrained their ability to shift rapidly around patchy or poor environmental conditions or degraded habitat.
Range of relative biomass predicted for Australian ecosystems in 2050 under climate change (vs. state in 2010).
| Group | GBR (NE) Australia | NW of Australia | SE of Australia |
|---|---|---|---|
| Biophysical indicators | |||
| Detritus | 0.91–1.13 | 0.84–1.07 | 0.97–1.28 |
| Primary producers | 1.47–1.65 | 0.97–1.04 | 0.95–1.2 |
| Plankton | 1.12–1.22 | 1.05–1.15 | 1.1–1.31 |
| Other pelagic invertebrates | 1.25–2.05 | 1.20–1.27 | 1.05–1.71 |
| Pelagic fish | 1.27–1.41 | 1.21–9.80 | 1.34–3.82 |
| Demersal fish | 0.9–0.92 | 0.65–0.81 | 0.68–0.93 |
| Benthic invertebrates | 1.05–1.12 | 0.77–0.88 | 0.91–0.96 |
| Top predators | 1.5–2.5 | 0.52–0.72 | 1.45–2.31 |
| Fisheries: economic indicators | |||
| Large-scale commercial sectors | 1.91–1.97 | 2.29–2.71 | 2.06–2.31 |
| Small-scale and recreational | 1.09–1.14 | 0.49–0.75 | 0.7–0.81 |
| Group | GBR (NE) Australia | NW of Australia | SE of Australia |
|---|---|---|---|
| Biophysical indicators | |||
| Detritus | 0.91–1.13 | 0.84–1.07 | 0.97–1.28 |
| Primary producers | 1.47–1.65 | 0.97–1.04 | 0.95–1.2 |
| Plankton | 1.12–1.22 | 1.05–1.15 | 1.1–1.31 |
| Other pelagic invertebrates | 1.25–2.05 | 1.20–1.27 | 1.05–1.71 |
| Pelagic fish | 1.27–1.41 | 1.21–9.80 | 1.34–3.82 |
| Demersal fish | 0.9–0.92 | 0.65–0.81 | 0.68–0.93 |
| Benthic invertebrates | 1.05–1.12 | 0.77–0.88 | 0.91–0.96 |
| Top predators | 1.5–2.5 | 0.52–0.72 | 1.45–2.31 |
| Fisheries: economic indicators | |||
| Large-scale commercial sectors | 1.91–1.97 | 2.29–2.71 | 2.06–2.31 |
| Small-scale and recreational | 1.09–1.14 | 0.49–0.75 | 0.7–0.81 |
Ranges represent the average values for the aggregate indicators (calculated over the entire model domain) for alternative model parametrizations.
Range of relative biomass predicted for Australian ecosystems in 2050 under climate change (vs. state in 2010).
| Group | GBR (NE) Australia | NW of Australia | SE of Australia |
|---|---|---|---|
| Biophysical indicators | |||
| Detritus | 0.91–1.13 | 0.84–1.07 | 0.97–1.28 |
| Primary producers | 1.47–1.65 | 0.97–1.04 | 0.95–1.2 |
| Plankton | 1.12–1.22 | 1.05–1.15 | 1.1–1.31 |
| Other pelagic invertebrates | 1.25–2.05 | 1.20–1.27 | 1.05–1.71 |
| Pelagic fish | 1.27–1.41 | 1.21–9.80 | 1.34–3.82 |
| Demersal fish | 0.9–0.92 | 0.65–0.81 | 0.68–0.93 |
| Benthic invertebrates | 1.05–1.12 | 0.77–0.88 | 0.91–0.96 |
| Top predators | 1.5–2.5 | 0.52–0.72 | 1.45–2.31 |
| Fisheries: economic indicators | |||
| Large-scale commercial sectors | 1.91–1.97 | 2.29–2.71 | 2.06–2.31 |
| Small-scale and recreational | 1.09–1.14 | 0.49–0.75 | 0.7–0.81 |
| Group | GBR (NE) Australia | NW of Australia | SE of Australia |
|---|---|---|---|
| Biophysical indicators | |||
| Detritus | 0.91–1.13 | 0.84–1.07 | 0.97–1.28 |
| Primary producers | 1.47–1.65 | 0.97–1.04 | 0.95–1.2 |
| Plankton | 1.12–1.22 | 1.05–1.15 | 1.1–1.31 |
| Other pelagic invertebrates | 1.25–2.05 | 1.20–1.27 | 1.05–1.71 |
| Pelagic fish | 1.27–1.41 | 1.21–9.80 | 1.34–3.82 |
| Demersal fish | 0.9–0.92 | 0.65–0.81 | 0.68–0.93 |
| Benthic invertebrates | 1.05–1.12 | 0.77–0.88 | 0.91–0.96 |
| Top predators | 1.5–2.5 | 0.52–0.72 | 1.45–2.31 |
| Fisheries: economic indicators | |||
| Large-scale commercial sectors | 1.91–1.97 | 2.29–2.71 | 2.06–2.31 |
| Small-scale and recreational | 1.09–1.14 | 0.49–0.75 | 0.7–0.81 |
Ranges represent the average values for the aggregate indicators (calculated over the entire model domain) for alternative model parametrizations.
In contrast to the consistency of the other groups, the relative biomass of benthic invertebrates and top predators responded differently in the two systems. In the NW, these two indicators declined—invertebrates by 12–23% and top predators by 28–48%, but they increased in the NE—invertebrates by 5–12% and top predators by 50–150%. In both systems, these results are driven by cascading effects of shifts in production, trophic interactions, and competition. The results also mask much skewed results across individual groups. For instance, despite the general level of change of invertebrates altogether, coral cover decreased by 60–85% (because of the mix of increased fragility resulting from acidification and more variable weather conditions). Among the species of conservation concern, the biomass of dugongs rose by 35–125%, because seagrass benefited from the slowing of calcifying epiphytic growth, whereas turtle populations collapsed (with many breeding beaches suffering inundation or storm disturbance). Moreover, although the gross biomass of demersal fish declined, this was largely concentrated among larger bodied or habitat-dependent groups, with the biomass of herbivorous and small demersal fish largely unaffected.
In the SE Australian model, on average (across alternative parametrizations of the model) the plankton, other pelagic invertebrates, and pelagic fish all again increased (by 10–31, 5–71, and 34–282%, respectively), whereas the demersal fish decreased by 7–32% (Table 3), all for similar reasons to those noted above. In this case, benthic invertebrates declined by 4–9% and the relative biomass of top predators increased by 45–131% (dominated by an increase in the great whales, which responded to increases in plankton production). These overall results again hide skewed and highly uncertain results for individual species (Figure 2), as well as significant spatial variability. One particularly interesting phenomenon was a predicted regime shift in the waters off eastern Bass Strait and Tasmania. This region of Australia has seen the largest changes in surface temperature over the past 60 years and is likely to see a further 2.6–4°C rise over the next 50 years under the emissions trajectory used in this analysis. Under this level of environmental change, Atlantis-SE predicts a change from a demersal and mesopelagically dominated system structure to one that is increasingly pelagically dominated (Figure 3). The shifts in nutrient patterns and thermal regime mean that there is an increase in both benthic and planktonic production by 20–50%, though the planktonic groups shift more towards picophytoplankton groups (whose contribution to production increases by 216%). There is a turnover in the types of benthic invertebrate (including some invading species) that dominate, moving to those that can cope with the changes in temperature, sources of production, detrital loads, and ocean acidification. The commercially valuable demersal fish initially increase (by 15–54%), because of more favourable temperatures and increases in some prey groups, but after 2040 they decline and are replaced by other demersal fish groups that feed on the new benthic invertebrate fauna coming to dominate the benthos. Similarly, squid and sharks initially increase (by 75–92 and 15–20%, respectively) but then also fall away after 2040 (the cephalopods in response to extended periods of environmental stress that outlasts their short life histories; the sharks because of a shift in prey mix, but also because they enter a suboptimal thermal regime). In contrast, the biomass of mesopelagics declines continuously, decreasing by 95%, whereas jellyfish and small pelagic forage fish increase by two- to fourfold or more (because they are either released from competition or, for the jellyfish, better able to cope with the new environmental conditions). Beyond 2060, the model becomes numerically unstable, because the parametrization (based on species composition and phenotypes in 2000) cannot continue to change under the changing conditions—model reparametrization would be required to reflect the level of biodiversity and evolutionary shifts that would have accumulated by that point.
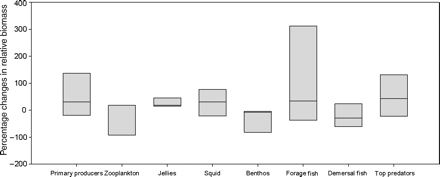
Range of results per functional group for Atlantis-SE climate change runs. The uncertainty bands indicate the range of results observed across all modelled groups across all simulations using alternative parametrizations. The median of each range is marked for reference.
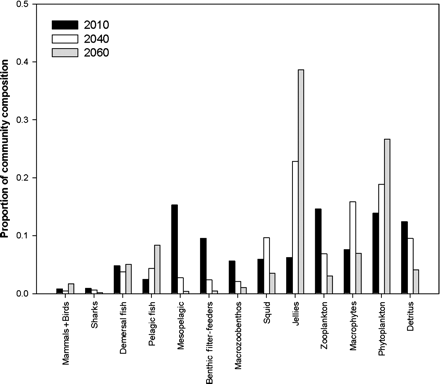
Regime shift predicted for the SE Australian ecosystem—proportional community composition in 2010, 2040, and 2060.
A comparison of these multifactor perturbation simulations with single-factor perturbation scenarios indicated that if factors affecting production (either via climate drivers or pollution) are considered in isolation, then results were within 10–15% of the values reached in the multifactor simulations (Table 4); although production and multifactor simulations diverge more in tropical systems than temperate systems. In contrast, considering the effects of acidification alone would result in a much more pessimistic view of future ecosystems—with values in the acidification-only runs 40% lower (on average) than in the multifactor scenarios. Once again, divergence was greater in tropical systems, where acidification could have quite devastating effects on habitat and associated assemblages.
Comparison of results from single and multifactor climate change perturbation scenarios for temperate (SE Australia) and tropical (NE and NW) Australian ecosystem models.
| Group | Temperate | Tropical | ||
|---|---|---|---|---|
| Production change only | Acidification only | Production only | Acidification only | |
| Biophysical indicators | ||||
| Detritus | 0.85–0.98 | 1.18–1.27 | 0.89–0.95 | 1.18–1.42 |
| Primary producers | 0.90–0.98 | 1.21–1.26 | 0.86–0.96 | 1.13–1.66 |
| Plankton | 0.94–0.98 | 1.25–1.61 | 0.91–0.96 | 1.17–1.51 |
| Other pelagic invertebrates | 0.96–0.98 | 1.26–1.37 | 0.97–0.98 | 1.32–1.71 |
| Pelagic fish | 0.83–0.98 | 1.33–2.51 | 1.15–1.22 | 1.14–1.31 |
| Demersal fish | 0.88–0.91 | 1.41–1.73 | 0.87–0.91 | 1.59–59.55 |
| Benthic invertebrates | 0.87–0.97 | 1.34–1.72 | 0.74–0.92 | 1.72–3.41 |
| Top predators | 0.91–0.96 | 1.48–1.82 | 0.51–0.68 | 1.71–1.97 |
| Group | Temperate | Tropical | ||
|---|---|---|---|---|
| Production change only | Acidification only | Production only | Acidification only | |
| Biophysical indicators | ||||
| Detritus | 0.85–0.98 | 1.18–1.27 | 0.89–0.95 | 1.18–1.42 |
| Primary producers | 0.90–0.98 | 1.21–1.26 | 0.86–0.96 | 1.13–1.66 |
| Plankton | 0.94–0.98 | 1.25–1.61 | 0.91–0.96 | 1.17–1.51 |
| Other pelagic invertebrates | 0.96–0.98 | 1.26–1.37 | 0.97–0.98 | 1.32–1.71 |
| Pelagic fish | 0.83–0.98 | 1.33–2.51 | 1.15–1.22 | 1.14–1.31 |
| Demersal fish | 0.88–0.91 | 1.41–1.73 | 0.87–0.91 | 1.59–59.55 |
| Benthic invertebrates | 0.87–0.97 | 1.34–1.72 | 0.74–0.92 | 1.72–3.41 |
| Top predators | 0.91–0.96 | 1.48–1.82 | 0.51–0.68 | 1.71–1.97 |
Values are the average from multifactor simulation/single-factor simulation.
Comparison of results from single and multifactor climate change perturbation scenarios for temperate (SE Australia) and tropical (NE and NW) Australian ecosystem models.
| Group | Temperate | Tropical | ||
|---|---|---|---|---|
| Production change only | Acidification only | Production only | Acidification only | |
| Biophysical indicators | ||||
| Detritus | 0.85–0.98 | 1.18–1.27 | 0.89–0.95 | 1.18–1.42 |
| Primary producers | 0.90–0.98 | 1.21–1.26 | 0.86–0.96 | 1.13–1.66 |
| Plankton | 0.94–0.98 | 1.25–1.61 | 0.91–0.96 | 1.17–1.51 |
| Other pelagic invertebrates | 0.96–0.98 | 1.26–1.37 | 0.97–0.98 | 1.32–1.71 |
| Pelagic fish | 0.83–0.98 | 1.33–2.51 | 1.15–1.22 | 1.14–1.31 |
| Demersal fish | 0.88–0.91 | 1.41–1.73 | 0.87–0.91 | 1.59–59.55 |
| Benthic invertebrates | 0.87–0.97 | 1.34–1.72 | 0.74–0.92 | 1.72–3.41 |
| Top predators | 0.91–0.96 | 1.48–1.82 | 0.51–0.68 | 1.71–1.97 |
| Group | Temperate | Tropical | ||
|---|---|---|---|---|
| Production change only | Acidification only | Production only | Acidification only | |
| Biophysical indicators | ||||
| Detritus | 0.85–0.98 | 1.18–1.27 | 0.89–0.95 | 1.18–1.42 |
| Primary producers | 0.90–0.98 | 1.21–1.26 | 0.86–0.96 | 1.13–1.66 |
| Plankton | 0.94–0.98 | 1.25–1.61 | 0.91–0.96 | 1.17–1.51 |
| Other pelagic invertebrates | 0.96–0.98 | 1.26–1.37 | 0.97–0.98 | 1.32–1.71 |
| Pelagic fish | 0.83–0.98 | 1.33–2.51 | 1.15–1.22 | 1.14–1.31 |
| Demersal fish | 0.88–0.91 | 1.41–1.73 | 0.87–0.91 | 1.59–59.55 |
| Benthic invertebrates | 0.87–0.97 | 1.34–1.72 | 0.74–0.92 | 1.72–3.41 |
| Top predators | 0.91–0.96 | 1.48–1.82 | 0.51–0.68 | 1.71–1.97 |
Values are the average from multifactor simulation/single-factor simulation.
The form and driving mechanisms for change in the different systems were largely consistent across model types; this engenders confidence in the robustness of the results. Where there were differences between the models, the trajectories and system dynamics differed in two main ways. First, Ecospace often predicted larger absolute responses across more individual groups than seen in either Atlantis or InVitro; this was typically because of stronger compensatory diet shifts in the later two models. However, as the regime shift outlined above indicates, once thresholds in these compensatory mechanisms were exceeded, Atlantis (and InVitro) can display sudden and substantial changes, as different mechanisms come to dominate. The second difference between the models was that Ecospace tended to display less interannual variation than evident in the other models. This is partly because of its longer time-step, which tended to have a smoothing effect on fluxes directed back to the coupled NPZD and because the forage groups in the Ecospace models were often more top-down controlled than in Atlantis and InVitro, which were more constrained by the productivity of lower trophic levels. In Atlantis and InVitro, large zooplankton, benthic invertebrates, and the youngest age classes of finfish all formed significant links between the upper and lower portions of the foodwebs. In contrast, squid and forage fish groups typically performed the linking role in Ecospace.
Economic outcomes
Economically, there are also quite uneven distribution of effects and benefits. In all the systems modelled, the large-scale commercial fisheries see a >90% increase in the relative value of their operations, because they have the socio-economic freedom to respond to new target mixes, distributions, and biomasses. In the NW and SE systems, the smaller scale (and recreational) fishers, who have less capacity for change, see a decline of 30–51%. For the GBR, this small-scale sector increased by 9–14%, but that is only if they were willing to accept small fish, because the trophy fish (large-bodied reef fish) they currently target are no longer a sustainable target (the catch of those groups declining by 22% or more). The increase in the value of the large-scale fisheries also hides differential results across sectors, with the prawn fishery suffering a 27–53% decline in value (mainly because of drastically reduced landings as the target stocks decline steeply).
Comparing dynamics across model types, the fisheries in Ecospace displayed less interannual variation and less variation between sectors than seen in the other models. The reason was the smoothing of interannual variation in the biophysical models, as discussed above, and because of the simpler representation of fisheries in the Ecospace models compared with the implementations of Atlantis and InVitro.
MMSY implications
The MMSY levels predicted for a 2060 system state was 27% lower for the tropical systems and 12% lower for SE of Australia, with the increase in biomass of the volatile pelagic groups unable to compensate fully for the losses of more stable demersal groups. This is consistent with what was found for other systems considered by Worm et al. (2009), with the majority of systems seeing a decline in MMSY under models forced with the most likely form of future environmental conditions (Figure 4). The only exceptions were the East Bering Sea and California Current, where there was very little, if any, change (<5% in both cases, because declines in some target species is offset by substantial increases in other, competing, commercially valuable groups).
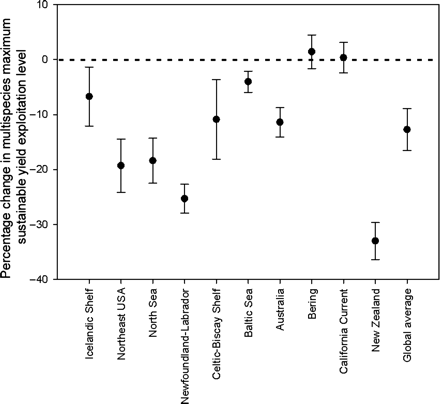
Change in the exploitation rate resulting in maximum multispecies sustainable yield (MMSY) under climate change. Dashed zero line given for reference.
Discussion
Climate change is expected to effect marine ecosystems via shifts in sea-level, temperature, incident solar radiation, pH, oxygen, stratification, surface winds, storm intensity and frequency, and changes in coastal run-off, upwelling, and current patterns (Hoegh-Guldberg, 2005; Hobday et al., 2006; Barange et al., 2010). Before 2010, there had been few published predictions of what those effects might be at an ecosystem level (i.e. simultaneously across all the interconnected aspects of an ecosystem from primary producers to top predators and the human users of the system). Most predictions had been based around changes in productivity or the distribution and abundance of single species or small closely linked groups (e.g. one to two forage species; Ito et al., 2010). One of the few exceptions is the global analysis by Cheung et al. (2009, 2010), which used a sophisticated extension to bioenvelope modelling to predict the aggregate effect of climate change on marine fisheries (Cheung et al., 2010) and biodiversity (Cheung et al., 2009). This approach still did not include trophic connections, but did result in similar levels of species invasions, local extinctions, and species turnover, as seen for the models presented here. However, although catch predictions by Cheung et al. (2010) are similar to those for the large-scale commercial fisheries for SE Australia, they underestimate commercial catch for the tropical systems, being closer to the predictions for the smaller sectors. Therefore, although there are some similarities, Cheung et al. (2010) do not give the sense of skewed outcomes seen in the current study. Neither do the predictions of Brown et al. (2009), which are focused on ecosystem and fisheries predictions for large-scale commercial fishing industries around Australia. Changes in pelagic productivity are similar in the analysis of Brown et al. (2009) and the current models, reflecting the fact that the current work was in part tuned to the same GCM-NPZ. The increase in the landings of the commercial fisheries predicted by Brown et al. (2009) is within 15% of the average change in overall landings predicted in this study. However, the skewed response across groups reported here is not reflected in the small changes to community composition recorded by Brown et al. (2009). However, to be fair, Brown et al. (2009) do clearly label it as a first step that does not include all the feedbacks and pressures that will be associated with climate change; therefore, it should not be surprising that it misses some of the skewness resulting from the omission of the multifactor combination of pressures included in the current study. For instance, turtles are predicted to increase in abundance by Brown et al. (2009), primarily driven by increases in productivity, whereas they are predicted to collapse in this study, largely because of the inundation of nesting beaches.
Despite the extra potential for non-linearity in the current study, at a system-level the shifts in the ecosystems seen here do largely tally with previous predictions. For instance, Boyd and Doney (2002), Bopp et al. (2005) and Morán et al. (2010) also predict that small phytoplankton groups coming to dominate. Furthermore, the increase in production in the SE and NW and the small decline in production along the GBR is in line with what is already being seen in SeaWiFS data (Behrenfeld et al., 2006)—though further data are needed to verify this finding, because the findings of Behrenfeld et al. (2006) could be the result of climate variability rather than climate change (Henson, et al., 2009). The large-scale decline in corals seen in the predictions here is in line with the predictions of Hoegh-Guldberg (1999), but is more pessimistic than the argument presented in Hughes et al. (2003), though even they caution that the effects of anthropogenically driven degradation of coral reefs and the associated habitat fragmentation have undermined reef resilience and increased their susceptibility to climate change.
Regime shifts, such as the one predicted for the SE Australian ecosystem in this study, have been an oft-mentioned potential outcome of climate change in many of the previous reviews (Brander, 2007, 2009; Barange et al., 2010). Examples from the North Atlantic (Beaugrand, 2004), Benguela (Shannon et al., 2008), and North Pacific (Hare and Mantua, 2000; Overland et al., 2008; Tian et al., 2008) illustrate how shifts in climatic properties could result in large-scale ecological restructuring. Similarly, switching between alternative foodweb pathways in Antarctica both seasonally and in high and low ice years (Atkinson et al., 2004) indicates how rapidly, but also how widespread, these effects can occur. Although regime shifts in the Baltic Sea (Möllmann et al., 2009) and Black Sea (Oguz et al., 2008) highlight how such shifts could be triggered, or at the very least exacerbated by, anthropogenic pressures, such as pollutant-based eutrophication, overfishing, and facilitation of bioinvasion. Regime shifts resulting directly from climate change are not expected to be ubiquitous, given that it is likely that at least some species that are lost from systems will be replaced by functional equivalents, meaning little, if any, net effect (Brander, 2009). However, because not all species will shift together, this will set the stage for potentially large-scale changes in community composition and new ecosystem structures (Hannah et al., 2005; Hoegh-Guldberg, 2005). Moreover, as seen in the predicted shift in the SE Australian ecosystem considered here, these changes can accrete from combinations of both slow shifts (e.g. resulting from generational-scale range expansions) and rapid turnover (e.g. either because of shifting migrations, growth or survival rates altering the mix and magnitude of species interactions) meaning that the pattern of change does not have be smooth or linear (Frank et al., 2005; Lovejoy and Hannah, 2005; Brander, 2007).
Perhaps the most consistent finding across all the models examined is that each system will have its own mix of winners and losers; something previously acknowledged as likely in reviews, such as those of Brander (2007, 2009). Although the current study suggests that pelagic systems might benefit and demersal systems decline, this generalization hides copious levels of species and group-level detail that is highly uncertain. This is reinforced by preliminary work (EAF, unpublished data) with a suite of 11 Atlantis and EwE models from major large marine ecosystems around the world, where there are strongly system-specific responses—some do match patterns for the Australian models discussed here, but for other systems all groups decline; in others the relative contributions of the demersal system increases, whereas the pelagic system does not. As with many things to do with ecosystems and their management, there will (generally) be no simple rules about how global change will affect marine ecosystems. Two exceptions to this are that it is already clear that there will be a mix of responses across groups and systems, with some doing better than others. Moreover, it is clear that the skewness in differential effects (and benefits), socially, economically, and ecologically will see the benefits rest with those groups (and sectors) with the greatest adaptive capacity. Again, this agrees with previous studies, such as the global economic analysis of the vulnerability of fisheries and nations dependent on them by Allison et al. (2009). The current study suggests that in Australian systems this seems to benefit primary producers and pelagic ecosystem components, which in some locations could see large-scale ecological restructuring. Economically, adaptive capacity seems to be with larger fisheries operators. As was found by Fulton et al. (2007), the smallest operators are most likely to suffer detrimental effects of system-level change. Although they might have lower cost structures, they are often more socially constrained and less able to shift as quickly or completely as larger operators—the reason is that the smaller operators have often made lifestyle choices or have family or other social commitments that mean that they cannot modify their behaviour as rapidly as “captains for hire” working for large companies (Baelde, 2001; Marshall et al., 2007). This pattern, of those with the least adaptive capacity being likely to be hit the hardest, is likely to be true across regions and nations (Brander, 2007, 2009; Allison et al., 2009), not just the sectors considered here.
Both the differential effects of climate change seen across the different ecosystem components and sectors simulated in the various models considered here, as well as the consideration of MMSY implications, indicate that modifications will have to be made to current management methods and reference points in response to changing marine ecosystems. Earlier reviews of potential climate effects on fisheries and management have already identified the confounding effect other human stressors can have on ecosystem responses to climate change (Hobday et al., 2006; Brander, 2009; Barange et al., 2010). Many have pointed to a need for more stringent management to reduce stress on ecosystems and give stocks more adaptive capacity (Roy and Pandolfi, 2005; Hobday et al., 2006; Brander, 2007). However, the differential effects observed here demonstrate that responses will have to be more nuanced than simply reducing pressure, if effective, egalitarian, and economically efficient management is to be assured. Eide and Heen (2002) found that the management strategy is as important, if not more so, than the climate scenario for determining the outcome in any one situation. Consequently, adaptive capacity must be considered, because it will influence the incentives of fishers and other stakeholders to comply with changing regulations (Fulton et al., 2011b). For instance, when recommending further cuts in fishing pressure in the fisheries considered in the MMSY analysis, it must be acknowledged that many might not have much scope for easily made reductions; cuts will likely see significant political opposition, because those same fisheries have already undergone extensive effort reductions over the past 10 years, having been restructured around more sustainable long-term effort levels (Worm et al., 2009). As with all instances of ecosystem-based management, the form of management required will have to vary with the system and cultures involved; recipe-book applications are unlikely to be productive (Fulton et al., 2011a). Although some generalities do exist, in the main there are sufficient differences in the form of the responses of social and ecological system components to climate and regulatory changes that the exact mix of levers will have to be tailored system-to-system.
The non-linear response of the modelled ecosystems considered here (e.g. the pattern of change 2010–2040–2060 seen in the Bass Strait region of the SE Australian model) indicates that not only should there be consideration of the mix of levers to use system-to-system, but also the form of management used will have to change as the system changes. Simply putting a static action in place will fail for most systems. This will further challenge existing management regimes. Currently, the best practice management methods are based on reference points, but this equilibrium concept will have to be updated for changing productivity (Brander, 2007), stock potential, and the new ecosystem structures. The latter two will prove particularly challenging, because they will be hard to predict, and noise in monitoring data will make the rate of signal detection around new states something that is relatively slow to determine relative to the cycle time of management regimes. One option is to maintain marine zoning that allows for the existence of reference areas. Quite apart from a conservation role, closed areas could become the new reference areas for judging the potential of new ecosystem structures and the magnitude of other anthropogenic effects outside these closures (Fulton et al., 2005). However, even with such a valuable role, effective spatial management will require a more fluid approach to the definition of marine zoning than is typically the case in places such as Australia, where acts of parliament are typically involved and they are therefore unlikely to be changed with any rapidity and are likely to fall out of step with the shifting distributions of marine species. Although a daunting task, the coordinated management of large spatial areas (at the regional-level scale that climate effects are evident) and across sectors is becoming a more common principle (Hannah et al., 2005; McDonald et al., 2006, 2008).
Although the results presented here have provided useful insights regarding system dynamics and management implications, they must be considered with caution. The reason is the issues identified in the description of the results for the Atlantis-SE model—namely the inability of a model with static parameters to capture fully the degree and kinds of change that will happen under the trending, but variable, pressure exerted by global change. Without explicit consideration of shifts in biodiversity, evolution, phenotypic plasticity, and other forms of adaptive capacity, these models can only provide constrained insights. Moreover, this is putting to one side the fact that the real world will present science (and society) with surprises that we cannot currently guess at (the “unknown unknowns” sensu Donald Rumsfeld 2002, then United States Secretary of Defense).
Supplementary material
The following supplementary material is available at the ICESJMS online version of this paper: the summaries and example formulations provided for the three end-to-end models are to afford readers with an appreciation of the models, their assumptions, and general forms. A summary of the biophysical and human components included in the specific implementations used as a basis for the Australian analyses presented in the main paper is provided in Table A1. Figure A1 is an example decision tree for animal agents in InVitro, and Figure A2 is a schematic diagram of agent types in different InVitro-NW and how the different submodels interconnect. TEPs stands for threatened, endangered and protected species (including whale sharks, marine mammals and seabirds). Bathymetry can directly influence all agent types; monitors can report (for governance or output purposes) on all other agent types; and turtle life histories are covered using multiple agent types.
Acknowledgements
I thank the CSIRO and both PICES and ICES for funding my participation in the International Symposium on Climate Change Effects on Fish and Fisheries held in Sendai, Japan, April 2010, which resulted in this paper being written. I also acknowledge funding from the Australian Fisheries Research and Development Corporation (FRDC) on behalf of the Australian Government and thank Miriana Sporcic, Scott Condie, and two anonymous reviewers for comments on an earlier draft on the manuscript. I am grateful to Cathy Bulman, Neil Gribble, Cameron Ainsworth, Alida Bundy, John Fields, Isaac Kaplan, Jason Link, Jodie Little, and Steve Mackinson for allowing me to use their Ecospace models as a basis for some of the work presented here. Last, I thank Bec Gorton, Gary Griffiths, and Penny Johnson for their technical support and fruitful discussions on the topic, and all the domain experts who have tolerated my seemingly inane questions while drawing these models together.

{kind=link}
{kind=link}
{kind=link}
{kind=link}