





Abstract
Understanding the spatial distribution and intensity of fishing activity is a prerequisite for estimating fishing impacts on seabed biota and habitats. Vessel monitoring system data provide information on fishing activity at large spatial scales. However, successive position records can be too infrequent to describe the complex movements fishing vessels make. High-frequency position data were collected to evaluate how polling frequency and the method of analysis influenced the estimates of fishing impact on the seabed and associated epifaunal communities. Comparisons of known positions with predictions from track interpolation revealed that the performance of interpolation depended on fleet behaviour. Descriptions and indicators of fishing intensity were influenced significantly by the analytical methods (track reconstruction, density of position records) and grid-cell resolution used for the analysis. These factors can lead to an underestimation of fishing impact on epifaunal communities. It is necessary to correct for such errors to quantify the effects of fishing on various ecosystem components and hence to inform ecosystem-based management. Polling at intervals of 30 min would provide a desirable compromise between achieving precise estimates of fishing impacts on the seabed and minimizing the cost of data collection and handling.Lambert, G. I., Hiddink, J. G., Hintzen, N. T., Hinz, H., Kaiser, M. J., Murray, L. G., and Jennings, S. 2012. Implications of using alternative methods of vessel monitoring system (VMS) data analysis to describe fishing activities and impacts. – ICES Journal of Marine Science, 69: 682–693.
Introduction
Ecosystem approaches to fisheries management often take account of the sustainability of fishing activities on the ecosystem, including seabed habitats (FAO, 2003; Sinclair and Valdimarsson, 2003). However, the capacity to assess ecosystem impacts will be influenced by the availability of detailed information on the location and intensity of bottom fishing activities. In Europe, vessel monitoring systems (VMSs) were introduced for fishery control and enforcement purposes, but are increasingly used to support the assessment of fishing activity and marine spatial planning (EC, 1997; Dinmore et al., 2003; Murawski et al., 2005; Mills et al., 2007). Given the original purpose of introducing such systems, the use of VMS data for research and impact assessment has some limitations. These include incomplete coverage of vessel activities, long durations between position records, and a lack of information on whether a vessel is actually fishing when the position is reported (Lee et al., 2010). At present, the last of these is inferred from a range of vessel speeds that relate to typical towing behaviour by different métiers (Dinmore et al., 2003). Generally, only vessels >15 m are monitored in Europe, and they typically transmit position records at intervals of 2 h (EC, 2009). Researchers therefore have to make assumptions and interpretations when using VMS data as the main source of information on fishing activities.
Even if methods can be developed to distinguish position records associated with fishing and non-fishing activities, it remains necessary to define methodologies and appropriate scales for reporting fishing activity and assessing its impact on the seabed. One option is to use the density of VMS position records as an index of fishing intensity. There are two ways in which this could be done, first using the density directly as an index and second by assigning an effort value to each VMS record and aggregating these values to a grid. These two approaches are probably most appropriate when describing activity in intensively fished areas and/or over large temporal and spatial scales, because they may underestimate intensity when applied to small areas and/or short periods through a lack of replication. For towed gears, a more complete assessment of the area fished can be made by interpolating between fishing position records.
The simplest approach for this is to use straight line (SL) interpolation between successive records, although this might underestimate track length and area fished when position records are infrequent (e.g. every 2 h), because most vessels fishing with towed bottom gears are not expected to maintain a straight course for such periods. An alternative method of track reconstruction is cubic Hermite spline interpolation (Hintzen et al., 2010), a technique that integrates information on vessel speed and heading at each polled position when computing the modelled vessel track. That method has been applied only to the Dutch flatfish beam trawl fishery to date, but it may be applicable to other towed gear fleets. Once tracks have been reconstructed, a grid of any given scale can be superimposed on the tracks and fishing activity can be reported per unit time per unit area and, when gear dimensions are known, as area swept per unit area per unit time. The grid scale of analysis used to accumulate fishing positions or to estimate fishing time or area has a notable effect on the interpretation of fishing distributions and the assumed impacts of fishing. For example, in any given region, the apparent extent of area without any fishing will decrease as the scale of the analysis is increased, with the result that total estimated fishing impact on the seabed will appear to be higher at larger scales of analysis (Dinmore et al., 2003; Piet and Quirijns, 2009). For these reasons, it is desirable to conduct an analysis of fishing intensity at the finest resolution possible, or at least at a resolution below which the distribution of activity is assumed to be random (Dinmore et al., 2003). The resolutions used to compute fishing intensity vary considerably: 1 km2 for the study of Nephrops trawling on benthic macrofauna or scallop fishing on habitat-forming species (Hinz et al., 2009; Lambert et al., 2011), 1 nautical mile2 (6.25 km2) for the study of trawling impacts on benthic infauna (Queirós et al., 2006; Reiss et al., 2009), 9 km2 for studying beam trawl impacts on benthic communities (Hiddink et al., 2006a), and 5 nautical mile2 for studying dredging impact on benthic communities (Hill et al., 1999; see also the review of Lee et al., 2010). However, these high-resolution analyses are vulnerable to deficiencies in the VMS data, because track reconstructions that do not capture the true path of a fishing vessel will lead to greater under- or overestimates of the intensity of effort at high resolution.
The grid scale of analysis is also important when fishing activity is used as a pressure indicator of the state of the environment. For example, EU directives on the Data Collection Framework (DCF) define standardized indicators of the integration of environmental protection requirements into the Common Fisheries Policy (COM, 2008; EC, 2008a, b). DCF indicators 5, 6, and 7 are the pressure indicators that quantify the distribution of fishing activities, the aggregation of fishing activities, and the non-impacted area of the seabed. Although they are intended to describe trends in pressure rather than absolute pressure values, the distribution of fishing is likely to be underestimated, the aggregation of fishing activities overestimated, and the non-impacted area of the seabed overestimated, if the indicators are calculated from individual position records rather than known or reconstructed tracks.
Detailed assessment of the performance of methods for estimating fishing activity is challenging when the real tracks of vessels are not known. In some cases, however, positions are recorded more frequently than the 2-h interval used as a standard in European fisheries. These high-frequency data allow interpolated positions based on 2-h data to be compared with known positions. The Isle of Man scallop fishery is one of these cases, position records of some vessels in the fishery having been recorded at 10–20-min intervals between November 2008 and October 2010. Here, we use these data to compare estimates of fishing activity determined with point-density and track-reconstruction methods at a range of spatial and temporal scales. The resulting estimates of fishing activity are then used to determine and compare the values of fishing pressure indicators for the distribution of fishing activities, the aggregation of fishing activities, and the non-impacted area of the seabed. Further, we assess the consequences of differences in estimated fishing activity distributions for the benthic community, based on an empirical analysis of the relationships between fishing pressure and the ecological status of the benthic community.
Material and methods
Interpolation of tracks
VMS position, time, speed, and anonymous vessel identification data for king scallop dredgers (DR) and queen scallop otter trawlers (OT) in the territorial waters of the Isle of Man for the period between November 2008 and October 2010 were provided by the Isle of Man Department of Environment, Food and Agriculture (DEFA). The data included Isle of Man and UK vessels ≥15 m, but also all vessels ≤15 m fishing in the 3-nautical mile zone, because all vessels fishing within that zone have to be fitted with VMS transmitters. Details on the species targeted were obtained by linking VMS and logbook using a unique vessel and date identifier (further detail in Murray et al., 2011). The data included high-poll-rate data resulting from control programmes led by DEFA and a specific request to provide additional such data for the present study. Before analysis, duplicate VMS records and records close to (within 1 km of) port were removed, along with erroneous position records allocated to land and selected records associated with fishing activity based on vessel speed (i.e. 43 383 records were removed; Lee et al., 2010). Initially, position records associated with fishing were identified by including all records with speeds of 0–6 knots to extract the tracks of individual vessels (Lee et al., 2010). However, based on direct observations of fishing vessels operating in these fisheries and an analysis of the frequency distribution of the speed, subsequent calculation of fishing intensity was restricted to speeds ≤3.4 knots (28 364 records). The cut-off speed of 3.4 knots was chosen based on the observation that data above that speed included several hundreds of VMS records that were situated in a known steaming corridor to the north of the Isle of Man. Any datapoints indicating a vessel speed between 0 (excluded) and 3.4 in the fishing area, i.e. not close to port, were considered to indicate fishing activity. The value zero was excluded to remove datapoints where vessels were anchored, but very low speeds were still included because vessels may slow down to empty dredges or to perform maintenance (Murray et al., 2011). All analyses were conducted using the VMStools package in R (Hintzen et al., 2012).
So-called real tracks, i.e. the exact paths followed by fishing vessels, were needed to parametrize and test the performance of different interpolation methods. Such tracks were assumed to be described by high-poll-rate data and compared with interpolations based on increasing time intervals of up to 2 h. In all, 86 and 12 vessel-days of such real tracks (position records at intervals <20 min, 4134 fishing pings) could be extracted from the VMS dataset for DR and OT dataset encompassing the period November 2008 to October 2010, respectively. Interpolation methods tested were the commonly used SL and cubic Hermit spline (cHs), the latter as developed by Hintzen et al. (2010). Succinctly, the cHs method uses information on vessel position, heading, and speed at times t and t+ 1 to define a trajectory. The combination of speed and heading are represented by vectors, and vector length is multiplied by a parameter fm that influences the curvature of the interpolations. Hintzen et al. (2010) describe two parameter optimization methods, one by minimizing the distance between the high-poll-frequency track (assuming SL interpolation between successive points) and the interpolated track, and the second by optimizing the length of the interpolated track to be as close as possible to that of the real track. Here, the former was adopted because the aim was to evaluate the accuracy of prediction of fishing locations.
Indicators of fishing pressure
Once the best models for predicting OT and DR fishing positions had been defined, the models were applied to the VMS dataset to build a dataset of interpolated positions. Any extraneous datapoints (speeds >3.4 knots) were removed as explained above. Two datasets were therefore used in the analyses: a cleaned dataset, hereafter referred to as the raw VMS dataset, and the dataset of interpolated positions. Interpolated positions were obtained by describing each interpolated track as a series of ten points at equal time intervals, the first and the last being the known positions at 2-h intervals and the intervening points the predicted positions. To compare quantitatively the spatial extent of fishing activity as estimated from raw VMS data or interpolated tracks, we calculated DCF pressure indicators 5, 6, and 7 (COM, 2008; EC, 2008a, b) and tested for the influence of spatial resolution on the derived estimates. Indicator 5 measures the area where fishing activity is reported for a defined bottom-fishing fleet and area. Indicator 6 measures the aggregation of fishing activities and is defined as the area (km2) in which 90% of the total activity for a defined bottom-fishing fleet in a defined area occurs. Indicator 7 measures the proportion of a defined area where no fishing activity is reported for a defined bottom-fishing fleet. The DCF and associated documentation recommends that the analysis be based on position records in 3 km × 3 km grid cells and to move from 2-h VMS position records to 30 min ones. The dataset of interpolated tracks was therefore also used to create a 30-min poll-rate dataset. The DCF indicators were then calculated for each of the three VMS datasets, i.e. raw VMS, interpolated tracks, and 30-min poll-rate datasets, along a grid cell size-gradient from 0.5 km × 0.5 km to 5 km × 5 km, at steps of 0.5 km × 0.5 km. Analyses were made on a log-transformed scale because were expected to be greater at the smaller spatial scales, so log-transforming the scale axis was expected to emphasize what was arising at smaller scales. As grid size was defined at constant kilometre units, longitude and latitude VMS position records were projected into the Universal Transverse Mercator (UTM) geographic coordinates system.
Fishing impact on epifaunal biomass
The relationship between fishing intensity and the biomass of sessile epifauna around the Isle of Man has been described before using the point summation of raw VMS data to estimate fishing intensity on a grid cell size of 1 km × 1 km (Lambert et al., 2011; using point summation as described below). However, other methods of estimating fishing intensity, e.g. using interpolated tracks, might be used for this purpose, so we considered the effects both of using different methods and their application at a range of spatial scales. The objective of the analysis was to assess how any differences in estimated distributions of fishing activity impacted on the relationship with the benthic community data and to assess the extent to which this affected the subsequent estimates of the impact of fishing on that community. Fishing intensity was estimated at 120 stations within 12 nautical miles of the Isle of Man where the seabed community was sampled during a 2008 survey of benthic habitats (Lambert et al., 2011). Fishing intensity was estimated by four methods at each station: (i) raw VMS data and grid, (ii) interpolated positions and grid, (iii) raw VMS data and Euclidean distance, and (iv) interpolated positions and Euclidean distance. Grid methods (i) and (ii) involved the placement of a grid over the whole area sampled and associating each benthic sample station with a grid cell and hence with the corresponding fishing intensity calculated within that cell. Euclidean distance methods (iii) and (iv) refer to cells defined by radial distance, i.e. based on the circular area surrounding each benthic sample station (Walter et al., 2007). The method places the sampled station at the centre of the area for which the fishing intensity is calculated. The Euclidean distance method seemed a priori more appropriate than the grid method to attribute a value of fishing intensity to a station, because a station rarely falls in the centre of a grid cell. However, most impact studies use the grid approach, because processed VMS data are often made available in that form, so we considered it important to quantify the differences between the two methods. The methods were applied at a series of spatial scales. Fishing intensity was calculated for the 12-month period before sampling because scallop fishing activities earlier would have determined the state of the seabed communities in August 2008. Only data for the period June 2007 to October 2008 were available. These data were not linked to information on vessel type, but analyses of data for the period between November 2008 and October 2010 allowed identification of the location of a Nephrops fishing ground to the west and southwest of the island. Therefore, the 15 stations in these areas were excluded from the analysis to focus solely on areas fished by king scallop DR during winter and queen scallop OT during summer. No other notable bottom-fishing métiers operate in the area.
For all four methods, fishing intensity was calculated by estimating the area swept by point summation. Point summation consists of summing the area swept attributed to each VMS point in a defined cell. The area swept was calculated by multiplying the time interval between positions, the fishing speed, and the breadth of the gear. Otter trawls were assumed to be 18.3 m wide, based on logbook data, and dredges from the UK were assumed to fish with the maximum number of dredges permitted by regulation, giving gear widths of 7.6 m inshore (0–3 nautical miles) and 12.16 m farther offshore (3–12 nautical miles; Lambert et al., 2011). These assumptions are likely to have resulted in overestimating the fishing activity following a precautionary approach to determining the impact of fishing, because not all vessels can deploy the maximum width of gear allowed (Murray et al., 2011). Estimates of swept-area associated with individual position records were then summed over a defined period or area for analysis. Fishing intensity is defined as the summed swept-area divided by the surface area of the cell (year–1) and was calculated for cell sizes ranging from ca. 0.5 km × 0.5 km to 5 km × 5 km for cells defined by the grid method corresponding to areas of 0.25–25 km2 for cells defined by radial distance, i.e. the Euclidean method. The latter led to overlap between cells, and fishing activity that belonged to several cells was independently attributed to each cell in which it fell, i.e. one fishing point could be attributed to two or more stations following the Euclidean distance method.
It was expected that, independent of methods, fishing intensity would vary with cell size in relation to the intensity of fishing activity around each station. Stations where fishing intensity is great at high resolution are likely to be attributed lower intensities when treated as part of larger areas, so intensity is averaged for those areas. The opposite phenomenon can be expected for lightly or non-impacted stations. These scale effects were examined before assessing how they influenced the apparent relationship between fishing and epifaunal biomass. The relationship between fishing intensity and scale was modelled for a range of quantiles from 0.1 to 0.9. Regressions on the 10th quantile (0.1) modelled changes for stations with the lowest fishing-intensity estimates along the scale gradient, and regressions on the 90th quantile (0.9) modelled changes for stations with the highest fishing-intensity estimates along the scale gradient. The slope and significance of the quantile regressions for each of the four fishing-intensity estimation methods described the changes in scale-dependent fishing-intensity estimates from the least-impacted (quantile 0.1) to the most-impacted (quantile 0.9) stations. The resolution gradient was log-transformed for reasons explained above.
As scale-dependent changes in fishing-intensity estimates at a station level were expected to impact the apparent relationship between fishing and epifaunal biomass, it was necessary to determine how the number of significant relationships between the biomass of sessile epifauna and fishing intensity varied with each fishing estimation method along the gradient of spatial resolution. Relationships were tested with linear quantile regressions (Koenker and Bassett, 1978; Cade and Noon, 2003; Lambert et al., 2011). Quantile regressions split the response data into quantile classes, so, in the present study, they are used to examine how the biomass of the benthic fauna is limited by a factor (fishing intensity) irrespective of the presence of other limiting factors (Blackburn et al., 1992; Cade and Noon, 2003). The 75th, 80th, 85th, 90th, and 95th quantiles of epifaunal biomass measurements of a range of functional and taxonomic groups were tested in relation to fishing intensity. Linear regressions were tested with and without log-transformation of fishing intensity. The functional groups were colonial vs. solitary, and hard vs. soft bodied species, and the taxonomic groups were Bivalvia, Balanidae, Cnidaria (including Hydrozoa and Anthozoa), Bryozoa, Porifera, and Ascidiacea (Lambert et al., 2011). For analysis of the impact on biomass, we included only stations where fishing intensity was ≤1 year–1, because fishing intensity was strongly right-skewed with very few stations (generally <5) fished more than once per year.
The impact of using a specific method and scale for quantitatively assessing the effect of fishing on sessile epifauna would further influence the predicted changes in epifaunal biomass at a regional scale, in this case Isle of Man territorial waters. Mortality as a consequence of dredging and otter trawling reduces the epifaunal biomass, and some of this “lost” biomass will be replaced by recruitment and growth. At a given station, the equilibrium relationship between biomass in the absence of fishing pressure (BI= 0) and fishing-impacted biomass (BI = F) can be expressed as BI=F = BI= 0e−F, where e–F is the proportion of BI= 0 remaining at fishing intensity F.
It was necessary too to assess how the predicted regional biomass of the sessile epifaunal community would vary when using different methods for calculating fishing intensity, with F based on raw VMS data on a grid, interpolated positions on a grid, raw VMS data and Euclidean distance, and interpolated positions and Euclidean distance, at resolutions of 0.5 km × 0.5 km to 5 km × 5 km. The value of e–F was estimated empirically from the slopes of the significant quantile regressions between sessile biomass and fishing intensity. Quantiles rather than means were used, because we were interested in the relative decline in biomass attributable to the limiting effect of fishing, i.e. the decrease in biomass caused by fishing, which is observed at the highest quantiles. The decrease in biomass was expressed as the predicted proportion of BI= 0 that remained when F increased from 0 to 1 year–1. The estimates of e–F and hence BI = F/BI= 0, the proportion of remaining biomass resulting from a fishing intensity of 1 year–1, obtained by the significant quantile regressions were averaged for each of four fishing-intensity estimation methods at each resolution.
Results
Interpolation of tracks
With cHs and SL interpolations, the spatial accuracy of interpolated tracks decreased as the time interval between VMS position records increased (Figure 1), and cHs better approximated the real tracks than SL at a given interval. For position records at 120-min intervals, both interpolation methods tracked real dredging (DR) tracks at comparable precision, whereas the distance between real tracks and interpolations improved by just 2% with cHs. The value of the optimized fm parameter declined rapidly towards zero (reflecting an SL track), when time intervals exceeded 90 min (time interval 120 min, fm= 0.0419; Figure 1). The real otter trawl (OT) tracks were better described by cHs interpolations than by SL, such that the distance to the real track at 120-min intervals was 17% smaller. At 120-min intervals, the reduction in the error of estimation of real track length by cHs was just 8% (ca. 0.5 km) for DR compared with 35% (ca. 2.6 km) for OT (Figure 1). Consequently, the results did not justify the use of a more complex model than SL for interpolating DR tracks, so subsequent estimation of fishing effort based on modelled tracks used that approach. However, cHs interpolations were judged to be more appropriate for interpolating OT tracks (fm = 0.1522).
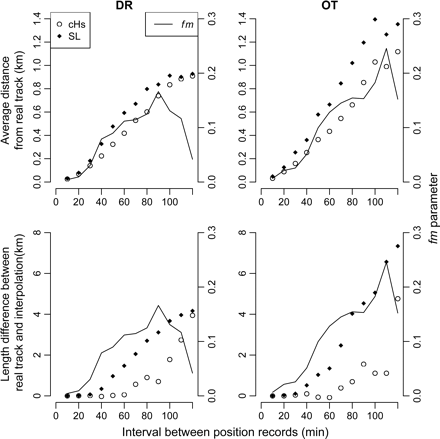
Distances between interpolated and known positions of tracks for DR and OT. Two interpolation scenarios are presented: cHs and SL. The distances are presented as a function of the interval between successive VMS position records (min). Upper panels represent the average distance (km) between real and interpolated tracks, and lower panels the difference in length estimates (km) between real and interpolated tracks. The optimized fm parameter shows how the interval increase affects the perceived behaviour of the fleet.
Indicators of fishing pressure
DCF indicators 5 and 6 showed that the area fished increased with the scale at which they were calculated and that DCF 7 mirrored these trends with a resultant decrease in the extent of the unfished area (Figure 2). The use of interpolated VMS data, or 30-min interval-interpolated VMS data, led to consistently higher estimates of DCF indicators 5 and 6 and lower estimates of DCF indicator 7 than when they were calculated with non-interpolated, raw VMS data (Figures 2 and 3). At the recommended resolution of 3 km × 3 km and poll intervals of 30 min, the area fished was just 9.5% smaller when DCF indicator 5 was calculated using raw VMS data than when it was calculated with interpolated VMS data. Aggregation of fishing activity, the focus of DCF indicator 6, was similarly 10.5% lower when computed with raw VMS data, which implied that the greater area covered by the interpolated data corresponded to areas fished recurrently. For DCF indicator 7, the area of the seabed not impacted by fishing decreased by ca. 18% when estimated from 30-min-interpolated rather than raw data (Figure 3).
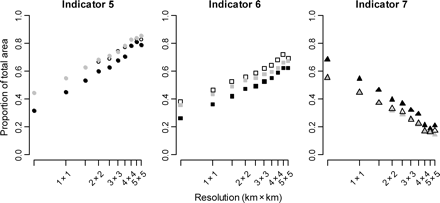
Estimated values of DCF indicators when calculated at a range of grid resolutions from high (0.5 km × 0.5 km) to low resolution (5 km × 5 km). Black symbols represent raw VMS data, grey ones data interpolated to 30-min intervals, and open symbols interpolated data. DCF indicator 5 represents the distribution of fishing activity, DCF indicator 6 the aggregation of fishing activity, and DCF indicator 7 the unfished area. All indicators are expressed as a proportion of the total area in Isle of Man territorial waters.
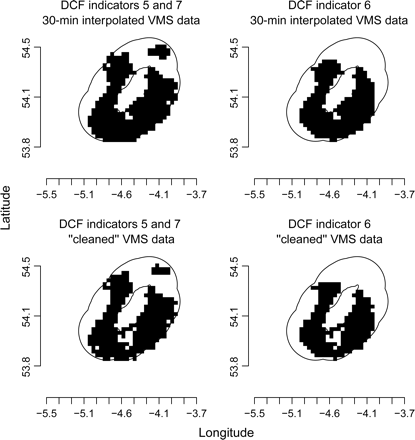
Maps of the annual distribution of the fishing activity used to calculate DCF indicators 5, 6, and 7, shown as 3 × 3 km grid cells. Upper panels are 30-min interpolated VMS data, and lower panels raw VMS data. Black cells are those impacted by scallop fishing gears, the white area that within the 12-nautical mile limit (black line), i.e. the non-impacted area of seabed.
Fishing intensity and the benthic community
The estimated fishing intensity at each station changed consistently with the resolution of the analysis. The coefficients of the slope of the quantile regressions between fishing intensity and scale were positive in the lowest range of quantiles (>0.1) and negative in the highest range (≥0.8). Hence, for stations that were least impacted, estimated fishing intensity increased with increasing resolution, whereas for the stations that were most impacted, estimated fishing intensity decreased with increasing resolution, with the steepest changes in estimated intensity at highest resolution (Figures 4 and 5). All four methods for estimating fishing intensity (raw VMS data with grid, interpolated positions with grid, raw VMS data with Euclidean distance, and interpolated positions with Euclidean distance) confirmed this trend, except the estimates based on interpolated positions, which did not decrease significantly with increasing resolution in the highest range of quantiles.
Changes in the estimated fishing intensity along the resolution gradient. Fishing intensity was estimated using the four methods shown on the panels. Changes are modelled separately at different quantile levels, from the 10th to the 90th, i.e. 0.1–0.9, showing the changes in estimates from the least to the most impacted stations. Slopes and significance per quantile are displayed in Figure 5.
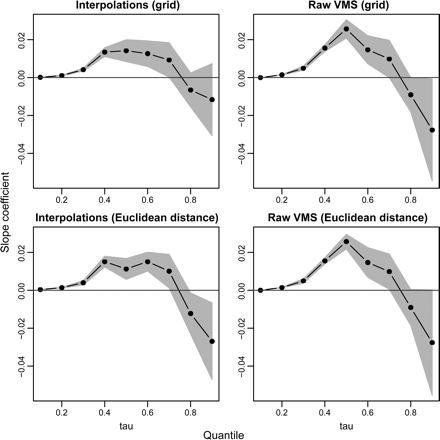
Slope coefficients of the quantile regressions for fishing intensity–scale as a function of the quantiles at which the regressions were tested. Fishing intensity was estimated using the four methods shown on the panels. The grey area represents the interpolated confidence interval of each effort coefficient estimate. The slope is significant when the confidence interval surrounding the estimate does not include zero. Quantile regressions for fishing intensity–scale are displayed in Figure 4.
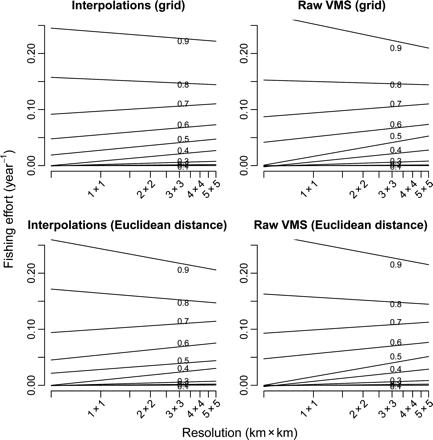
Despite the changes in estimated fishing intensity among the different methods and scales of resolution, there was broad qualitative evidence that, in general, increasing fishing intensity reduced sessile epifauna biomass (Figure 6). However, the significance of the results differed according to the methods by and scales at which fishing intensity was calculated. Across the gradient of grid-cell resolution, the number of significant quantile regressions peaked at a spatial scale between 2 km × 2 km and 4 km × 4 km (Figure 7). In this range, i.e. 2 km × 2 km and 4 km × 4 km, the relationships between fishing intensity, as estimated from interpolated tracks and Euclidean distance, and epifaunal community biomass were the most consistent. Grid-based methods and raw VMS data gave the least consistent results across the different spatial resolutions used to estimate fishing intensity.
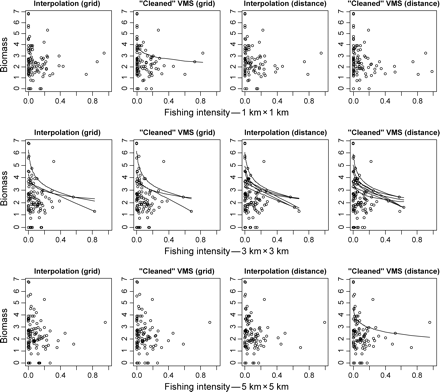
Relationships between fourth-root-transformed biomass of emergent epifauna and fishing-intensity measurements. The biomass was fourth-root transformed consistently with the methods described in Lambert et al. (2011). Each pair of panels represents fishing intensity calculated at a different resolution (described in the x-axes), and each row represents four scenarios where fishing intensity at the sampled stations is calculated on a grid or based on Euclidean distance and where VMS data are used either raw or interpolated. Only significant linear quantile regressions on fishing intensity with or without log-transformation are shown.
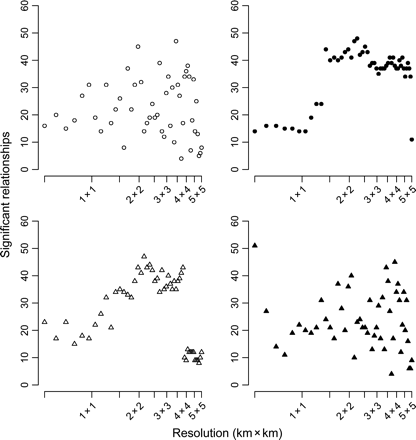
The number of significant relationships between biomass and fishing intensity as a function of the resolution chosen to calculate fishing intensity. Fishing intensity was measured using four different methods. Circles represent fishing intensities calculated on a grid, and triangles fishing intensities calculated from Euclidean distance, with open symbols representing interpolated data (left column) and black-filled symbols raw VMS data (right column). The number of significant relationships derives from univariate quantile regressions between biomass at different taxonomic and functional levels and fishing intensity gradients (see text).
The estimated proportion of epifaunal biomass that remained after being impacted by fishing varied between 0 and 0.5 depending on the method and resolution used to estimate fishing intensity. A value of 1.0 for the remaining proportion of biomass arose as a result of non-significant relationships between total biomass of epifauna and fishing intensity for certain combinations of method and resolution (Figure 8). Grid-based outputs were those most affected by spatial resolution as highlighted by the variation in the remaining proportion of epifaunal biomass, and Euclidean-based outputs were more stable. Excluding the resolutions at which no significant quantile regression had been observed, the estimated proportion of the remaining epifaunal biomass after a fishing impact of 1 year–1 decreased as scale increased for both Euclidean-distance-based methods, i.e. higher spatial resolutions significantly decreased the perceived effect of fishing activities. The fit of the linear model that showed the decrease in predicted remaining biomass with lower resolutions of estimation of fishing effort was better when fishing effort was calculated from interpolated tracks than from raw VMS data (respectively, d.f. = 30, p < 0.001, r2= 0.49, and d.f. = 30, p = 0.037, r2= 0.11; Figure 8).
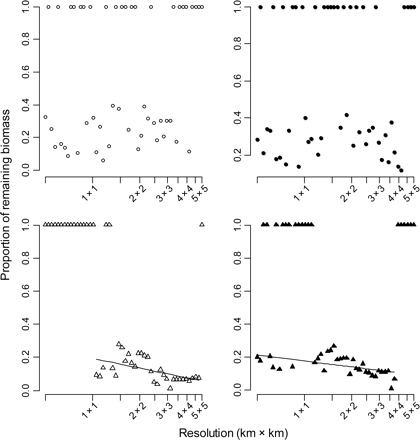
The proportion of biomass remaining after a fishing impact of 1 year–1 as a function of the spatial resolution at which the fishing intensity is calculated. The proportion 1 reflects non-significant quantile regressions. Fishing intensity was measured with four different methods. Circles represent fishing intensities calculated on a grid, and triangles fishing intensities calculated from Euclidean distance, with open symbols representing interpolated data (left column) and black-filled symbols raw VMS data (right column). Significant linear regressions are also displayed (excluding the proportion of remaining biomass equal to 1, see text for statistics).
Discussion
Many approaches can be used to analyse VMS data. The results here show that different approaches have both strengths and weaknesses depending on the scale at which the analysis is undertaken and the scientific questions addressed. The results also illustrate the general challenges of using relatively infrequent position records, which are considered sufficient to meet many fisheries-enforcement needs, to describe the distribution and wider ecosystem impacts of fishing activities. Unusually, we were able to quantify the performance of methods for estimating fishing intensity from infrequent VMS records because we had access to more-frequent position records.
SL and cHs interpolation led to different results for the different fleets, and both Isle of Man fleets behaved differently from the Dutch beam trawl fleet, for which the method was developed and applied by Hintzen et al. (2010). The Dutch fishery deploys large vessels that trawl over relatively homogeneous substrata, with minimum delay between hauling and resetting gear. The Isle of Man scallop fishery targets restricted grounds in daylight. The high polling frequency VMS data and information from fishers demonstrated that Isle of Man vessels tend to go back and forth over the same ground, leading to complex track patterns in fishing grounds that may only be 3–5 km in the longest dimension. Within the Isle of Man fleet, the fishing tracks of scallop DR and queen scallop OT have different characteristics. DR can make sharp 180° turns to dredge directly alongside their previous tracks, whereas OT usually require larger turning circles (LGM, NTH, pers. obs.). These differences in behaviour explain why SL fitted better the tracks of scallop DR, whereas cHs was a better method for OT. Of course, the estimates of the model parameters could have been refined if more high-frequency data had been available and if those data could have been split into a training dataset and a test dataset.
Although the results show that the availability of more-frequent position records would describe fishing tracks better, increased polling comes at an increased cost. The European Commission has recommended 30 min as a suitable compromise between providing better information for calculating fishing pressure indicators and minimizing the cost of additional polling. However, the results here show that there is always an error in the spatial location of tracks for the Isle of Man scallop fishery at even the lowest poll frequencies. Notwithstanding the fact that accurately interpolating fishing tracks from 1 or 2 h VMS data is possible for fisheries such as the Dutch flatfish beam trawl fishery (Hintzen et al., 2010), the results here demonstrate that this rule cannot be applied across different fleets without first considering behavioural patterns specific to that fleet. Reducing the polling frequency from 2 to 1 h would reduce the error by a factor of ∼2 for DR and by 3 for OT and improve estimates of track length and hence of wider ecosystem impacts. In practice, the results show that the different characteristics of different fisheries mean that no single polling rate will provide the best compromise between cost and the resolution of fishing activity. Regulators, ideally informed by research that takes account of non-regulatory uses of VMS data, need to consider whether it would be practical for different rates to be used by different fleets and how such fleets would be defined. Alternatively, random high-frequency polling could provide the necessary data to compute the extent of uncertainty in modelled estimates of fishing intensity and allow fine-tuning of the applied methods, as appropriate.
More-frequent information on fishing positions, as obtained by more-frequent polling or track reconstruction, will yield point density estimates that better capture the true distribution of fishing activity. In the current case, the difference in the values of DCF indicators showed that the true distribution of fishing activities was underestimated by ca. 10% when tracks were not interpolated. This means that areas considered not impacted using raw VMS data based on a 2-h polling interval had in reality been impacted to some extent. Errors in the interpolation of tracks might also result in some areas being wrongly identified as impacted. These potential biases are especially relevant given that the first pass of a trawl or dredge has proportionately greater impact on the benthic community than subsequent passes over the same area (Hiddink et al., 2006b). The difference between the values of indicators based on interpolation and raw VMS data decreased as the scale of analysis increased. For Isle of Man territorial waters, the difference between unfished areas estimated from raw VMS data and from interpolated data decreased from ca. 500 to ca. 50 km2 with increasing grid-cell resolution. This difference arises because the finer-resolution analyses reflected better the heterogeneity in fishing effort distribution. Failure to work at a resolution that adequately described this heterogeneity can lead to overestimates of the magnitude of fishing impacts, irrespective of the method used to calculate fishing intensity (Mills et al., 2007; Piet and Quirijns, 2009).
Different grid-cell resolutions and methods of fishing-intensity estimation changed the significance of the relationships between fishing intensity and epifaunal biomass. The most significant relationships were associated with spatial resolutions of 2 km × 2 km to 4 km × 4 km. This may seem inconsistent with other studies of fishing impacts that attempted to use the smallest possible grid cell to quantify fishing pressure, often on the basis that the distribution of fishing was assumed to be random at that scale (Rijnsdorp et al., 1998; Dinmore et al., 2003; Reiss et al., 2009). There are two potential reasons for this discrepancy. First, observed changes in the benthic fauna of an impacted site may also depend on the effects of fishing impacts in the area surrounding that location. These impacts may, for example, define how fast animals can recolonize the station through direct immigration or larval recruitment. Such effects will be accounted for if fishing intensity is estimated at larger a spatial scale. Second, spatial resolutions of 2 km × 2 km to 4 km × 4 km that lead to the most significant relationships between biomass and fishing intensity were based on a single year's VMS data. If there is year-to-year variation in fishing intensity, but recovery from fishing impacts takes place over longer periods, then aggregated annual data over larger scales may provide a more reliable picture of the distribution of fishing activities and a better assessment of their cumulative impacts on the benthic community. To avoid increased patchiness within cells, the selection of grid-cell resolution is a trade-off between the need to aggregate sufficient records to obtain a reliable index of intensity over an appropriate period and the need to ensure that the effect of patchiness is not overlooked.
The scale of analysis also influences the predicted biomass remaining after fishing. The estimated fishing intensity at stations subject to the greatest impacts at small scales will tend to be influenced by less impact in surrounding areas, whereas the opposite phenomenon can arise at the stations that are least impacted. The decrease in the remaining biomass of sessile organisms predicted when the scale of analysis increases may therefore reflect the decrease in estimated fishing intensity at stations highly impacted at greatest resolution. This is because highly impacted stations at which very low biomass was observed will be attributed a lower estimate of fishing intensity, which would create a steeper decrease in biomass along the fishing gradient.
Preferred methods of fishing-intensity estimation depend on the objectives of the study and the spatial and temporal resolution of the available VMS data. The results here broadly support the conclusions of Lee et al. (2010) who suggested that point-density methods on cleaned VMS data were suitable for providing information on patterns of fishing activity on large scales of space and time, but that track-based methods would be more appropriate when attempting to ascertain the spatial impact of vessels using mobile gears over shorter periods of time. Given the diversity of fisheries and fishing intensities, spatial and temporal trade-offs will vary among fisheries, so no singular solution will exist and a compromise would have to be adopted if the same resolution were to be applied to multiple fisheries. For mapping fishing activity, 3 km × 3 km has been considered a reasonable compromise (Mills et al., 2007). However, a resolution greater than 3 km × 3 km would be needed to describe the effects of specific disturbance events on seabed habitats, given the typical resolution of biological sampling. For instance, the time since the last fishing disturbance event will affect community composition at a sampling station, and such a station will typically cover a much smaller area than 3 km × 3 km (Tillin et al., 2006).
In general, the results here show that more-frequent position records and finer analytical resolutions support more-accurate and informed assessment of fishing activity and its subsequent impact on benthic biota. Although current data collected in Europe have provided new insights into the distribution of fishing activity on smaller scales of time and space than was hitherto feasible, the limitations of such data are apparent when they are used to describe impacts at local scales over short periods. Clearly, greater polling frequencies provide more information on real tracks and overcome the need for more complex and fleet-specific interpolation methods, but the results here do not suggest that there would be a fleet-specific optimum. Rather, the existing 2-h intervals seem to be too long, so any achievable reduction in polling frequency would be desirable. A reduction to 30 min would appear to be an appropriate intermediate target and would provide significant benefits when VMS data are used for mapping activity and assessing fishing impacts, but the long-term target would be to increase polling frequency to a rate appropriate for the behaviour and fishing grounds of a specific fleet.
Acknowledgements
We thank the UK Department of Environment, Food and Rural Affairs (Defra Project M1001) for funding the research, and the Isle of Man Department of Environment, Food and Agriculture for permission to use the data and photographs from their habitat mapping survey conducted in August 2008. Collection and quantification of biological data were carried out with the assistance of many Bangor University students, the efforts of all of whom are gratefully acknowledged. Finally, we thank two anonymous reviewers for their valued comments on the submitted text.
References
Author notes
Handling editor: Andrew Payne

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}