





Abstract
Seasonal time-series of temperature and salinity around Iceland have been collected, with repeat observations at stations, since the start of the 1970s. After fairly cold years during the early 1990s, a reversal occurred during the mid-1990s and the period from 1996 to 2010 was one with warmer, more saline water in shelf seas around Iceland. Temperatures and salinities have been frequently above the long-term mean in the waters to the south and west of the country, and these conditions have also influenced the shelf area north of the country. Marked changes have also been observed in the distribution of many fish species during this warm period. Southern commercial species have extended farther north (e.g. haddock, monkfish, mackerel), a northern species is retreating (capelin), rare species and vagrants have been observed more frequently (e.g. greater fork beard, blue antimora, snake pipefish, sea lamprey, Ray's bream), and 31 species, from both shelf and oceanic waters, have been recorded for the first time since 1996. The most obvious explanation for these changes in the ecosystem has to be the warming of 1–2°C in the waters south and west of Iceland during the past 15 years.Valdimarsson, H., Astthorsson, O. S., and Palsson, J. 2012. Hydrographic variability in Icelandic waters during recent decades and related changes in distribution of some fish species. – ICES Journal of Marine Science, 69: 816–825.
Introduction
Icelandic waters are in a frontal zone where the warmer water of the North Atlantic Current and the Irminger Current flowing northwards to the Nordic Seas meet the south-flowing colder waters of the East Greenland Current and the East Icelandic Current (Figure 1). A small part of the Irminger Current flowing north along the western shelf of Iceland then flows northeastwards towards the shelf area north of Iceland as the North Icelandic Irminger Current (Stefánsson, 1962; Malmberg and Kristmannsson, 1992; Valdimarsson and Malmberg, 1999; Malmberg and Valdimarsson, 2003). Observations have shown large interannual and decadal hydrographic variability on the northern shelf, and cooling and freshening periods have been noted as part of larger scale climatic variations such as the Great Salinity Anomaly (Dickson et al., 1988) and have had a marked influence on the living conditions for various biota (Jakobsson, 1992; Astthorsson et al., 2007). Then, after the mid 1990s, observations showed warming and salinification in the waters south and west of Iceland (Malmberg and Valdimarsson, 2003).
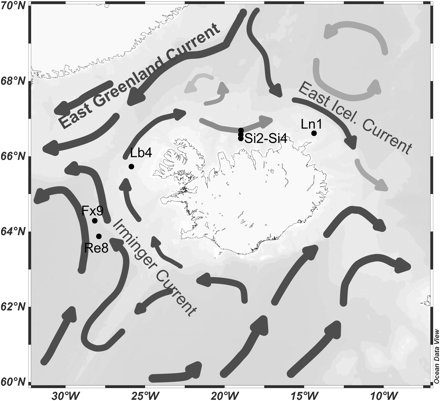
Main ocean currents and hydrographic sections in Icelandic waters. The stations referred to in the article (Fx9, Lb4, Si2–Si4, Ln1) are noted with black dots. Based on Valdimarsson and Malmberg (1999) and Hunegnaw et al. (2009).
During a continuous warm period in the waters around Iceland since 1996, seawater temperature off the south and west shelves has increased by ∼1–2°C. Further, the temperatures during the decade 2000–2010 have been the highest of the observation period and there are no indications that a reversal is about to occur. During this same period, extensive shifts in the distribution and increase in the abundance of many fish stocks have also taken place. This pertains to several commercially exploited stocks, some fairly common species that are not exploited, southern vagrants, and finally many new species have been recorded during this recent warm period (Valdimarsson et al., 2005; Astthorsson and Palsson, 2006; Astthorsson et al., 2007, 2010; Palsson 2007, 2008, 2009, 2010). In this paper, we summarize the findings from previous studies of the changes in the distribution of fish species, which hitherto have mostly been reported in Icelandic and discuss them in relation to recent hydrographic variability.
Material and methods
Seasonal observations in Icelandic waters have been undertaken since 1970 along hydrographic sections and at fixed stations (Figure 1). Other data from repeated stations are available from more or less annual observations in spring or summer from as early as 1948 (Stefánsson, 1962). Until 1988, observations of temperature and salinity were obtained by water bottles, and thereafter the measurements were performed using CTDs. Data presented in the time-series shown here are mainly from the period covered by the seasonal data, and observations from within a 3-week window have been used principally. Local warming is small compared with interannual variability, especially in winter and spring, because the stratification by then has not normally developed markedly. Data from the water-bottle period have been interpolated and then depth-averaged before time-series were constructed. Here, information is presented from two stations and a mean of three stations from sections in the waters to the southwest, northwest, and north of Iceland.
The results presented on the distribution and abundance of the exploited and other relatively common fish species are mainly based on material sampled during routine bottom-trawl surveys of fish stocks in Icelandic waters. Information on rare fish and new fish species similarly originates from research cruises, with additional information from Icelandic fishers, who have routinely brought fish that they have not seen before or have awoken their interest to the Marine Research Institute. These fish have been caught using various kinds of fishing gears (Table 1).
Fish species recorded for the first time and locations within the Icelandic EEZ, 1996–2010.
| Years of record | English name | Latin name | Number of stations | Number of specimens | Area | Depth (m) | Gear | Notes |
|---|---|---|---|---|---|---|---|---|
| 1996 | – | Pseudoscopelus altipinnis | 1 | 1 | S, W | Deep water | M | Detailed position and depth recorded not available |
| 1996, 1999, 2004, 2006, 2007 | Black devil anglerfish | Melanocetus johnsoni | 5 | 5 | S, W | 700–1 400 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 1996–2004 | Blue shark | Prionace glauca | >100 | >1 000 | S | <50 | L | Record possibly connected to gear type and/or research outside traditional grounds (see main text) |
| 1997 | – | Macroparalepis affinis | 1 | 1 | NW | 300 | B | Northernmost record to our knowledge (Jonsson and Palsson, 2006) |
| 1997, 1998, 2001, 2002, 2006, 2009 | – | Chaunax suttkusi | 9 | 9 | S, W, NW | 300–900 | B | Distribution recently expanding northwards (Quigley et al., 1996; Quero et al., 1998) |
| 1998 | Legless searsid | Platytroctes apus | 2 | 2 | S | 600–800 | M | Nearest previous records from Denmark Strait (Jonsson and Palsson, 2006) |
| 1998 | Atlantic bonito | Sarda sarda | 1 | 1 | NW | <10 | G | Caught in coastal water (Arnarfjord) |
| 1998, 1999, 2001 | Violet cuskeel | Brotulotaenia crassa | 3 | 3 | S | 200–700 | M | Sporadic records in the eastern North Atlantic (Nielsen, 1986b) |
| 1998 | – | Neonesthes capensis | 1 | 1 | S | 700 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1998 | Headlight fish | Diaphus effulgens | 1 | 1 | S | 500 | B | Nearest previous record west of Ireland (Hulley, 1984) |
| 1998 | Brownsnout spookfish | Dolichopteryx longipes | 1 | 1 | S | 700–800 | P | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1999–2010 | Flounder | Platichthys flesus | >20 | >100 | S, W, NW, N | 0–50 | G, D, Et | See text |
| 1999, 2001, 2003 | Pink sabretooth | Evermannella balbo | 4 | 4 | S | 500–1 000 | M | Nearest previous records from southern Irminger Sea (MRI, unpublished data) |
| 2000 | – | Lamprogrammus shcherbachevi | 1 | 1 | S | 50–100 | B | Only three previous records from North Atlantic (Nielsen and Schwarzhans, 2000) |
| 2000, 2001, 2002, 2003 | – | Lycodes terraenovae | 9 | 9 | S, W | 1 100–2 400 | B, A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2000, 2006 | Deepscale dory | Grammicolepis brachiusculus | 2 | 2 | S, NW | 300–400 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 2001, 2004 | Palebelly searsid | Barbantus curvifrons | 3 | 3 | W | 500–800 | M | Known from the northern Mid Atlantic Ridge (Gushchin and Kukuev, 1981) |
| 2001 | – | Haplophryne mollis | 1 | 1 | S | 500 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 2001 | – | Poromitra megalops | 2 | 10 | S | 500–800 | M | See text |
| 2002 | Carapine grenadier | Coryphaenoides carapinus | 1 | 1 | S | 1700 | A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2002, 2008 | Sailfin dory | Zenopsis conchifera | 5 | 6 | S, W | 75–400 | D, B | See text |
| 2004 | John dory | Zeus faber | 1 | 1 | S | 75–100 | B | See text |
| 2006 | Allis shad | Alosa alosa | 1 | 1 | S | 150–250 | B | Known from European coastal water (Whitehead, 1984) |
| 2006 | Blackmouth catshark | Galeus melastomus | 1 | 1 | S | 200–300 | B | Known in eastern North Atlantic from Senegal to Norway (Quéro, 1984) |
| 2006 | – | Linophryne maderensis | 1 | 1 | S | 700–800 | P | Previously only known from off the coast of Madeira (Bertelsen, 1984) |
| 2006 | Longspined bullhead | Taurulus bubalis | 5 | W | 0–5 | Pn | See text | |
| 2007 | Narrow-necked oceanic eel | Derichthys serpentines | 1 | 1 | S | 300–1 000 | M | Northernmost record to our knowledge (Bauchot and Saldanha, 1986) |
| 2007 | Rendezvous fish | Polymetme corythaeola | 1 | 1 | S | 400 | B | Northernmost record to our knowledge (Badcock, 1984) |
| 2007 | Atlantic batfish | Dibranchus atlanticus | 1 | 1 | S | 550–700 | B | Northernmost record to our knowledge (Quero et al., 1998) |
| 2007 | – | Linophryne pennibarbata | 1 | 1 | S | 1 400 | B | Northernmost record to our knowledge (Bertelsen, 1980) |
| 2008, 2009 | Four-spot megrim | Lepidorhombus boscii | 5 | 8 | S, W | 150–250 | B | See text |
| Years of record | English name | Latin name | Number of stations | Number of specimens | Area | Depth (m) | Gear | Notes |
|---|---|---|---|---|---|---|---|---|
| 1996 | – | Pseudoscopelus altipinnis | 1 | 1 | S, W | Deep water | M | Detailed position and depth recorded not available |
| 1996, 1999, 2004, 2006, 2007 | Black devil anglerfish | Melanocetus johnsoni | 5 | 5 | S, W | 700–1 400 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 1996–2004 | Blue shark | Prionace glauca | >100 | >1 000 | S | <50 | L | Record possibly connected to gear type and/or research outside traditional grounds (see main text) |
| 1997 | – | Macroparalepis affinis | 1 | 1 | NW | 300 | B | Northernmost record to our knowledge (Jonsson and Palsson, 2006) |
| 1997, 1998, 2001, 2002, 2006, 2009 | – | Chaunax suttkusi | 9 | 9 | S, W, NW | 300–900 | B | Distribution recently expanding northwards (Quigley et al., 1996; Quero et al., 1998) |
| 1998 | Legless searsid | Platytroctes apus | 2 | 2 | S | 600–800 | M | Nearest previous records from Denmark Strait (Jonsson and Palsson, 2006) |
| 1998 | Atlantic bonito | Sarda sarda | 1 | 1 | NW | <10 | G | Caught in coastal water (Arnarfjord) |
| 1998, 1999, 2001 | Violet cuskeel | Brotulotaenia crassa | 3 | 3 | S | 200–700 | M | Sporadic records in the eastern North Atlantic (Nielsen, 1986b) |
| 1998 | – | Neonesthes capensis | 1 | 1 | S | 700 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1998 | Headlight fish | Diaphus effulgens | 1 | 1 | S | 500 | B | Nearest previous record west of Ireland (Hulley, 1984) |
| 1998 | Brownsnout spookfish | Dolichopteryx longipes | 1 | 1 | S | 700–800 | P | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1999–2010 | Flounder | Platichthys flesus | >20 | >100 | S, W, NW, N | 0–50 | G, D, Et | See text |
| 1999, 2001, 2003 | Pink sabretooth | Evermannella balbo | 4 | 4 | S | 500–1 000 | M | Nearest previous records from southern Irminger Sea (MRI, unpublished data) |
| 2000 | – | Lamprogrammus shcherbachevi | 1 | 1 | S | 50–100 | B | Only three previous records from North Atlantic (Nielsen and Schwarzhans, 2000) |
| 2000, 2001, 2002, 2003 | – | Lycodes terraenovae | 9 | 9 | S, W | 1 100–2 400 | B, A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2000, 2006 | Deepscale dory | Grammicolepis brachiusculus | 2 | 2 | S, NW | 300–400 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 2001, 2004 | Palebelly searsid | Barbantus curvifrons | 3 | 3 | W | 500–800 | M | Known from the northern Mid Atlantic Ridge (Gushchin and Kukuev, 1981) |
| 2001 | – | Haplophryne mollis | 1 | 1 | S | 500 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 2001 | – | Poromitra megalops | 2 | 10 | S | 500–800 | M | See text |
| 2002 | Carapine grenadier | Coryphaenoides carapinus | 1 | 1 | S | 1700 | A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2002, 2008 | Sailfin dory | Zenopsis conchifera | 5 | 6 | S, W | 75–400 | D, B | See text |
| 2004 | John dory | Zeus faber | 1 | 1 | S | 75–100 | B | See text |
| 2006 | Allis shad | Alosa alosa | 1 | 1 | S | 150–250 | B | Known from European coastal water (Whitehead, 1984) |
| 2006 | Blackmouth catshark | Galeus melastomus | 1 | 1 | S | 200–300 | B | Known in eastern North Atlantic from Senegal to Norway (Quéro, 1984) |
| 2006 | – | Linophryne maderensis | 1 | 1 | S | 700–800 | P | Previously only known from off the coast of Madeira (Bertelsen, 1984) |
| 2006 | Longspined bullhead | Taurulus bubalis | 5 | W | 0–5 | Pn | See text | |
| 2007 | Narrow-necked oceanic eel | Derichthys serpentines | 1 | 1 | S | 300–1 000 | M | Northernmost record to our knowledge (Bauchot and Saldanha, 1986) |
| 2007 | Rendezvous fish | Polymetme corythaeola | 1 | 1 | S | 400 | B | Northernmost record to our knowledge (Badcock, 1984) |
| 2007 | Atlantic batfish | Dibranchus atlanticus | 1 | 1 | S | 550–700 | B | Northernmost record to our knowledge (Quero et al., 1998) |
| 2007 | – | Linophryne pennibarbata | 1 | 1 | S | 1 400 | B | Northernmost record to our knowledge (Bertelsen, 1980) |
| 2008, 2009 | Four-spot megrim | Lepidorhombus boscii | 5 | 8 | S, W | 150–250 | B | See text |
No new species were recorded in 2003, 2005, 2009, or 2010. The areas of occurrence south (S), west (W), northwest (NW), north (N), and east (E) are based on Saemundsson (1949). The abbreviations for fishing gears are midwater trawl (M), longline (L), bottom trawl (B), gillnet (G), Danish seine (D), Agassiz trawl (A), eel trap (Et), and pushnet (Pn). For some of the lesser known species, no English names exist.
Fish species recorded for the first time and locations within the Icelandic EEZ, 1996–2010.
| Years of record | English name | Latin name | Number of stations | Number of specimens | Area | Depth (m) | Gear | Notes |
|---|---|---|---|---|---|---|---|---|
| 1996 | – | Pseudoscopelus altipinnis | 1 | 1 | S, W | Deep water | M | Detailed position and depth recorded not available |
| 1996, 1999, 2004, 2006, 2007 | Black devil anglerfish | Melanocetus johnsoni | 5 | 5 | S, W | 700–1 400 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 1996–2004 | Blue shark | Prionace glauca | >100 | >1 000 | S | <50 | L | Record possibly connected to gear type and/or research outside traditional grounds (see main text) |
| 1997 | – | Macroparalepis affinis | 1 | 1 | NW | 300 | B | Northernmost record to our knowledge (Jonsson and Palsson, 2006) |
| 1997, 1998, 2001, 2002, 2006, 2009 | – | Chaunax suttkusi | 9 | 9 | S, W, NW | 300–900 | B | Distribution recently expanding northwards (Quigley et al., 1996; Quero et al., 1998) |
| 1998 | Legless searsid | Platytroctes apus | 2 | 2 | S | 600–800 | M | Nearest previous records from Denmark Strait (Jonsson and Palsson, 2006) |
| 1998 | Atlantic bonito | Sarda sarda | 1 | 1 | NW | <10 | G | Caught in coastal water (Arnarfjord) |
| 1998, 1999, 2001 | Violet cuskeel | Brotulotaenia crassa | 3 | 3 | S | 200–700 | M | Sporadic records in the eastern North Atlantic (Nielsen, 1986b) |
| 1998 | – | Neonesthes capensis | 1 | 1 | S | 700 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1998 | Headlight fish | Diaphus effulgens | 1 | 1 | S | 500 | B | Nearest previous record west of Ireland (Hulley, 1984) |
| 1998 | Brownsnout spookfish | Dolichopteryx longipes | 1 | 1 | S | 700–800 | P | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1999–2010 | Flounder | Platichthys flesus | >20 | >100 | S, W, NW, N | 0–50 | G, D, Et | See text |
| 1999, 2001, 2003 | Pink sabretooth | Evermannella balbo | 4 | 4 | S | 500–1 000 | M | Nearest previous records from southern Irminger Sea (MRI, unpublished data) |
| 2000 | – | Lamprogrammus shcherbachevi | 1 | 1 | S | 50–100 | B | Only three previous records from North Atlantic (Nielsen and Schwarzhans, 2000) |
| 2000, 2001, 2002, 2003 | – | Lycodes terraenovae | 9 | 9 | S, W | 1 100–2 400 | B, A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2000, 2006 | Deepscale dory | Grammicolepis brachiusculus | 2 | 2 | S, NW | 300–400 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 2001, 2004 | Palebelly searsid | Barbantus curvifrons | 3 | 3 | W | 500–800 | M | Known from the northern Mid Atlantic Ridge (Gushchin and Kukuev, 1981) |
| 2001 | – | Haplophryne mollis | 1 | 1 | S | 500 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 2001 | – | Poromitra megalops | 2 | 10 | S | 500–800 | M | See text |
| 2002 | Carapine grenadier | Coryphaenoides carapinus | 1 | 1 | S | 1700 | A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2002, 2008 | Sailfin dory | Zenopsis conchifera | 5 | 6 | S, W | 75–400 | D, B | See text |
| 2004 | John dory | Zeus faber | 1 | 1 | S | 75–100 | B | See text |
| 2006 | Allis shad | Alosa alosa | 1 | 1 | S | 150–250 | B | Known from European coastal water (Whitehead, 1984) |
| 2006 | Blackmouth catshark | Galeus melastomus | 1 | 1 | S | 200–300 | B | Known in eastern North Atlantic from Senegal to Norway (Quéro, 1984) |
| 2006 | – | Linophryne maderensis | 1 | 1 | S | 700–800 | P | Previously only known from off the coast of Madeira (Bertelsen, 1984) |
| 2006 | Longspined bullhead | Taurulus bubalis | 5 | W | 0–5 | Pn | See text | |
| 2007 | Narrow-necked oceanic eel | Derichthys serpentines | 1 | 1 | S | 300–1 000 | M | Northernmost record to our knowledge (Bauchot and Saldanha, 1986) |
| 2007 | Rendezvous fish | Polymetme corythaeola | 1 | 1 | S | 400 | B | Northernmost record to our knowledge (Badcock, 1984) |
| 2007 | Atlantic batfish | Dibranchus atlanticus | 1 | 1 | S | 550–700 | B | Northernmost record to our knowledge (Quero et al., 1998) |
| 2007 | – | Linophryne pennibarbata | 1 | 1 | S | 1 400 | B | Northernmost record to our knowledge (Bertelsen, 1980) |
| 2008, 2009 | Four-spot megrim | Lepidorhombus boscii | 5 | 8 | S, W | 150–250 | B | See text |
| Years of record | English name | Latin name | Number of stations | Number of specimens | Area | Depth (m) | Gear | Notes |
|---|---|---|---|---|---|---|---|---|
| 1996 | – | Pseudoscopelus altipinnis | 1 | 1 | S, W | Deep water | M | Detailed position and depth recorded not available |
| 1996, 1999, 2004, 2006, 2007 | Black devil anglerfish | Melanocetus johnsoni | 5 | 5 | S, W | 700–1 400 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 1996–2004 | Blue shark | Prionace glauca | >100 | >1 000 | S | <50 | L | Record possibly connected to gear type and/or research outside traditional grounds (see main text) |
| 1997 | – | Macroparalepis affinis | 1 | 1 | NW | 300 | B | Northernmost record to our knowledge (Jonsson and Palsson, 2006) |
| 1997, 1998, 2001, 2002, 2006, 2009 | – | Chaunax suttkusi | 9 | 9 | S, W, NW | 300–900 | B | Distribution recently expanding northwards (Quigley et al., 1996; Quero et al., 1998) |
| 1998 | Legless searsid | Platytroctes apus | 2 | 2 | S | 600–800 | M | Nearest previous records from Denmark Strait (Jonsson and Palsson, 2006) |
| 1998 | Atlantic bonito | Sarda sarda | 1 | 1 | NW | <10 | G | Caught in coastal water (Arnarfjord) |
| 1998, 1999, 2001 | Violet cuskeel | Brotulotaenia crassa | 3 | 3 | S | 200–700 | M | Sporadic records in the eastern North Atlantic (Nielsen, 1986b) |
| 1998 | – | Neonesthes capensis | 1 | 1 | S | 700 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1998 | Headlight fish | Diaphus effulgens | 1 | 1 | S | 500 | B | Nearest previous record west of Ireland (Hulley, 1984) |
| 1998 | Brownsnout spookfish | Dolichopteryx longipes | 1 | 1 | S | 700–800 | P | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 1999–2010 | Flounder | Platichthys flesus | >20 | >100 | S, W, NW, N | 0–50 | G, D, Et | See text |
| 1999, 2001, 2003 | Pink sabretooth | Evermannella balbo | 4 | 4 | S | 500–1 000 | M | Nearest previous records from southern Irminger Sea (MRI, unpublished data) |
| 2000 | – | Lamprogrammus shcherbachevi | 1 | 1 | S | 50–100 | B | Only three previous records from North Atlantic (Nielsen and Schwarzhans, 2000) |
| 2000, 2001, 2002, 2003 | – | Lycodes terraenovae | 9 | 9 | S, W | 1 100–2 400 | B, A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2000, 2006 | Deepscale dory | Grammicolepis brachiusculus | 2 | 2 | S, NW | 300–400 | B | Northernmost record to our knowledge (Astthorsson and Palsson, 2006) |
| 2001, 2004 | Palebelly searsid | Barbantus curvifrons | 3 | 3 | W | 500–800 | M | Known from the northern Mid Atlantic Ridge (Gushchin and Kukuev, 1981) |
| 2001 | – | Haplophryne mollis | 1 | 1 | S | 500 | M | Nearest previous records from Denmark Strait (MRI, unpublished data) |
| 2001 | – | Poromitra megalops | 2 | 10 | S | 500–800 | M | See text |
| 2002 | Carapine grenadier | Coryphaenoides carapinus | 1 | 1 | S | 1700 | A | Record possibly connected to gear type and/or research outside traditional grounds |
| 2002, 2008 | Sailfin dory | Zenopsis conchifera | 5 | 6 | S, W | 75–400 | D, B | See text |
| 2004 | John dory | Zeus faber | 1 | 1 | S | 75–100 | B | See text |
| 2006 | Allis shad | Alosa alosa | 1 | 1 | S | 150–250 | B | Known from European coastal water (Whitehead, 1984) |
| 2006 | Blackmouth catshark | Galeus melastomus | 1 | 1 | S | 200–300 | B | Known in eastern North Atlantic from Senegal to Norway (Quéro, 1984) |
| 2006 | – | Linophryne maderensis | 1 | 1 | S | 700–800 | P | Previously only known from off the coast of Madeira (Bertelsen, 1984) |
| 2006 | Longspined bullhead | Taurulus bubalis | 5 | W | 0–5 | Pn | See text | |
| 2007 | Narrow-necked oceanic eel | Derichthys serpentines | 1 | 1 | S | 300–1 000 | M | Northernmost record to our knowledge (Bauchot and Saldanha, 1986) |
| 2007 | Rendezvous fish | Polymetme corythaeola | 1 | 1 | S | 400 | B | Northernmost record to our knowledge (Badcock, 1984) |
| 2007 | Atlantic batfish | Dibranchus atlanticus | 1 | 1 | S | 550–700 | B | Northernmost record to our knowledge (Quero et al., 1998) |
| 2007 | – | Linophryne pennibarbata | 1 | 1 | S | 1 400 | B | Northernmost record to our knowledge (Bertelsen, 1980) |
| 2008, 2009 | Four-spot megrim | Lepidorhombus boscii | 5 | 8 | S, W | 150–250 | B | See text |
No new species were recorded in 2003, 2005, 2009, or 2010. The areas of occurrence south (S), west (W), northwest (NW), north (N), and east (E) are based on Saemundsson (1949). The abbreviations for fishing gears are midwater trawl (M), longline (L), bottom trawl (B), gillnet (G), Danish seine (D), Agassiz trawl (A), eel trap (Et), and pushnet (Pn). For some of the lesser known species, no English names exist.
Results and discussion
Hydrographic variability
The Faxafloi section west of Iceland stretches to the west over the Icelandic shelf and into the outer core of the Irminger Current, which lies over the shelf break and slope. The last station on this section (Fx9; Figure 1) is located in this core and shows the character of the Atlantic water that flows upstream to the Denmark Strait. This station has been occupied from May 1983, when the Faxafloi section replaced the earlier Reykjanes sections. Earlier observations exist from station Re8 (Figure 1) on the Reykjanes section (20 nautical miles farther south, 7 miles farther east), which has a similar bottom depth as Fx9. Calibration profiles taken within a couple of hours interval have shown similar temperatures and salinities at both locations. Hence, the time-series from Re8 and Fx9 have been combined into an extended series for station Fx9 that has been used to represent the northern Irminger Sea (Mortensen and Valdimarsson, 1999; Reverdin et al., 1999).
The extended Fx9 station was also used to represent one of the Atlantic inflow branches into the Arctic Mediterranean and to compare with the simulated Subpolar Gyre Index. Hátún et al. (2005) showed high correlation between the SGI and the salinity at Fx9 shifted back 2 years to adjust for advective delays. Long-term variations of temperature and salinity at this station are similar to those at deeper stations on the standard sections south and southeast of Iceland, so this station can be said to represent stations in the Atlantic water south of the Greenland–Scotland Ridge.
The salinity and temperature at Fx9 (Figure 2) increased rather rapidly in 1996 after a period of low values at the start of the 1990s and earlier. Both temperature and salinity reached peak values in 1998 and then remained high. Similar changes have been observed at all sections from the southeast, south, and west of Iceland (ICES, 2010a).
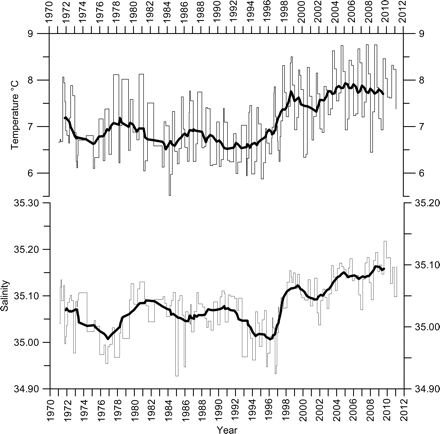
Seasonal temperature and salinity (0–200 m) at station Fx9 on the Faxafloi section (for location, see Figure 1). The bold line depicts the 3-year running averages.
The Látrabjarg section lies farther north on the western shelf and extends westwards into the Denmark Strait. Station Lb4 (Figure 1) is located on this section on the shelf in Atlantic water flowing northwards. The temperature of the upper layers in summer at Lb4 has been increasing since the middle of the 1990s (Figure 3), and this represents the trends for the shelf west and south of Iceland in that season. Temperature in August 2010 was the highest for 35 years (Figure 3), and the five cases in which the temperature has been above 10°C have all been during the years 2000–2010. The uppermost 50-m layer in summer is also greatly influenced by local variability in wind mixing and insolation, adding to the effect of advection of warmer water.
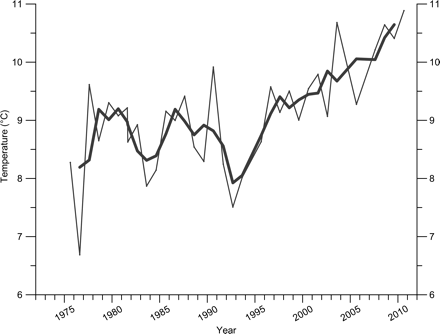
Surface temperature (mean from the surface to 50 m) at station Lb4 on the Latrabjarg section during the period 1975–2010 (for location, see Figure 1). The bold line depicts the 3-year running averages.
The observed changes in temperature and salinity to the south and west have also, to a certain degree, been advected to the area north of Iceland, as is demonstrated by February observations at the Siglunes section. Figure 4 shows the depth average of temperature and salinity between 50 and 150 m for three stations, Si2–Si4 (Figure 1), which span a north–south range of 20 nautical miles. These stations and depth intervals are located in the core of the Atlantic inflow, which reaches the northern area with the North Icelandic Irminger Current (Stefánsson, 1962; Valdimarsson and Malmberg, 1999). By selecting this depth interval, we avoid to some degree the noise induced by occasional surface melt water in the area.
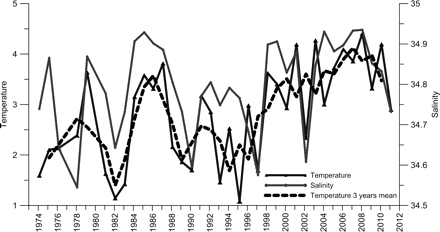
Temperature (°C) and salinity (mean 50–150 m) during winter at stations Si2–Si4 on the Siglunes section during the period 1970–2011 (for location, see Figure 1).
At stations Si2–Si4, the average temperature in winter since 1996 has been above the long-term mean for the observation period (1974–2011) during 13 of 15 winters. The two winters when temperature was below the mean were ones when sea ice drifted into the northern area, resulting in short-term cooling and freshening. These winter temperatures can be looked at as indicators of lower limits, and it can be speculated that the more frequent higher winter values have resulted in fish species from warmer areas having a better chance of survival on the northern shelf. Similar warming and spreading of more saline water has also been observed on shelf stations along the path of the inflowing Atlantic water on the northern shelf farther to the east (Anon., 2008). Hence, during the period 1973–2010, when summer (August) observations are available at Langanes section (station Ln1; Figure 1), five of seven of the highest observed temperatures were in the years 2000–2010.
Changes in water mass characteristics in Icelandic waters in the 1960s and 1970s have mainly been related to freshening and cooling in the Nordic Seas. After the mid-1990s, the observed warming and salinification in the waters south and west of Iceland were suggested to be similar to the warm hydrographic conditions that prevailed during the period 1920–1960 (Malmberg and Valdimarsson, 2003). This has now turned out to be the case for the waters around Iceland and larger areas of the northern North Atlantic and Nordic Seas. This has been reported in many papers over the past decade or so, and the increased heat and salinity in the Iceland Basin and in the Irminger Sea has been related to larger scale changes in dynamics and hydrography of the Subpolar Gyre (Berch, 2002; Hakkinen and Rhines, 2004; Hátún et al., 2005). Hakkinen and Rhines (2009) studied drifter data, revealing a large-scale shift in surface currents in the early 2000s. Further, Hakkinen et al. (2011) related the variable amount of subtropical water penetrating polewards into the Subpolar Gyre to windstress curl, whereas other local forcing, such as Greenland tip jet events and recurring blocking patterns, were also considered to be having an effect.
Changes in fish distribution
Exploited stocks
With respect to commercially exploited fish stocks in Icelandic waters, many species have shifted their distribution during recent warm period. Among these are capelin (Mallotus villosus), haddock (Melanogrammus aeglefinus), blue whiting (Micromesistius poutassou), saithe (Pollachius virens), Icelandic summer-spawning herring (Clupea harengus), oceanic redfish (Sebastes mentella), mackerel (Scomber scombrus), monkfish (Lophius piscatorius), and lemon sole (Microstomus kitt; Gudmundsdottir and Sigurdsson, 2004; Valdimarsson et al., 2005; Astthorsson and Palsson, 2006; Astthorsson, 2007; Astthorsson et al., 2007).
Of these species, capelin is the only Arctic one, and during the recent warming, it has retreated northwestwards from its usual nursery and feeding grounds to the north of Iceland to colder waters over the East Greenland shelf (Palsson et al., 2009; Anon., 2011). At the same time, recruitment has been low, and this has led to a decrease in stock size. Whether poor recruitment (during the period 2001–2009 all years except 2005 and 2009 have been poor) and/or change in larval drift is also related to the warming conditions is not clear.
Of the other species that have demonstrated a distributional shift, all are of southern origin and, except summer-spawning herring, they have all been caught in greater quantities than before in the waters north of Iceland during recent years. The most marked changes with respect to these exploited stocks of the southern origin relate to haddock, monkfish, and mackerel.
Haddock in the waters south of Iceland have been considered to be at the northern limit of their distribution. Between 1998 and 2003, all year classes except that of 2001 were above average (the one from 2003 was exceptionally large), and therefore the stock size increased and, probably because of higher temperature, the area of distribution extended farther north along the west coast and onto the northern shelf (Anon., 2007; Astthorsson et al., 2007). During the most recent years, the haddock stock has again decreased somewhat because of lower recruitment, but it is still widely distributed on the northern shelf (Anon., 2011). Two other southern gadoids, Norway pout (Trisopterus esmarkii) and whiting (Merlangius merlangus), which are subjected to only limited exploitation, have also increased in abundance and extended their distribution north in recent years (Valdimarsson et al., 2005; Astthorsson and Palsson, 2006; Bjornsson et al., 2007).
Monkfish was until recently mainly confined to limited areas in the warm water off the south and east coasts of Iceland (Jonsson and Palsson, 2006). Until about 2000, they were mainly caught as a bycatch in a lobster fishery, and from 1965 to 2000, the average annual catch was ∼700 t. Since 1998, large cohorts have recruited almost annually, which is in great contrast to at least 15 years before that. This improved reproductive success has been attributed to extended nursery areas becoming available owing to the higher temperature and salinity (Solmundsson et al., 2010a). Similarly, during the past decade, the distribution of this species has extended along the whole of the south and west coasts and onto the northern shelf (Figure 5). This recent increase in stock size has led to a targeted fishery on monkfish, with an average catch of ∼3000 t during the past 5 years (Anon., 2011).

The spatial distribution of monkfish in the Icelandic Groundfish Survey, 1985–2009 (after Solmundsson et al., 2010b).
In the eastern North Atlantic, mackerel are distributed from the Azores and Cape Verde Islands, into the Mediterranean and Black Seas, and north to northern Norway and even the White Sea (Iversen, 2004). Mackerel have been considered a vagrant in Icelandic waters during the past 100 years, and records of occurrence are mainly confined to three periods: several early records at the turn of the 20th century; the years between 1925 and 1955; and at the turn of the 21st century (Astthorsson et al., 2010, in press). The latter two periods conform to the warming that took place in the North Atlantic during the early part of the 20th century (e.g. Drinkwater, 2006) and the warming in recent years (Hughes et al., 2010). Most of the past records of mackerel in Icelandic waters report only a single, or a few, fish in any given year (Astthorsson et al., 2010, in press), but during the past 3 years, mackerel have been found in the area in unprecedented numbers and become widely distributed almost all around Iceland (Figure 6). This sudden shift in distribution appears to have arisen without a marked increase in stock size (ICES, 2010b), and therefore it is most likely related to changes in the environment. In recent years, the zooplankton biomass in the Norwegian Sea has been steadily decreasing (ICES, 2010c), and Loeng et al. (2009) pointed out that this was possibly related to overgrazing of the large pelagic stocks inhabiting the area. In Icelandic waters, there are no signs of such a steady decrease in zooplankton (MRI, unpublished data), so possibly the westward shift in the distribution of mackerel may be driven by the combined effect of higher temperatures and favourable feeding conditions in the waters around Iceland. The shift in the mackerel distribution has led to the development of a direct fishery inside the Icelandic Exclusive Economic Zone (EEZ), which from 2008 to 2010, amounted to more than 100 000t annually.
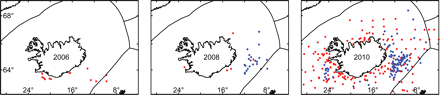
Locations of mackerel samples analysed by the Marine Research Institute from Icelandic and nearby waters in during 2006, 2008, and 2010. Red dots are samples from Icelandic research vessels and blue dots are from the Icelandic pelagic fishing fleet.
Species of low abundance
Some smaller southern fish stocks in Icelandic waters, which have been found mainly off the south coast, have also been caught in increasing numbers in the shelf water north of Iceland during recent warm years (Astthorsson and Palsson, 2006). Examples of these species are the greater forkbeard (Phycis blennoides), blue antimora (Antimora rostrata), and snake pipefish (Entelurus aequoreus). The snake pipefish is of particular interest because it seems to have been increasing in abundance and extending its distribution over wide areas in the northeastern North Atlantic (Kirby et al., 2006; Lindley et al., 2006; van Damme and Couperus, 2006; Fleischer et al., 2007; Harris et al., 2007; Rusyaev et al., 2007). According to Jonsson and Palsson (2006), snake pipefish are distributed in the Northeast Atlantic Ocean from the coasts of the Azores and Portugal north to the middle of the west coast of Norway. In Icelandic waters, it has until recently been found mainly off the southwest coast. In 2001, snake pipefish were recorded for the first time for more than a decade in Icelandic waters when a single specimen was found off the west coast (Astthorsson and Palsson, 2006). Additional records off the south and west coasts were further registered in 2002, 2003, 2004, and 2007. In autumn 2004, snake pipefish were also recorded from off the north coast, and in 2005, 2006, and 2007, additional records were registered from that area (Astthorsson and Palsson, 2008). These last records demonstrate that snake pipefish can now be considered to be distributed all around Iceland.
Vagrants
Several southern vagrant fish species have been recorded more frequently in Icelandic waters in recent years. Among these are, for example, twaite shad (Alosa fallax), sea lamprey (Petromyzon marinus), garfish (Belone belone), and ocean sunfish (Mola mola). Figure 7 shows the occurrence of these species in the period 1992–2009. Before 1992, they had not, except for lamprey, been observed for more than a decade (Jonsson and Palsson, 2006). Following the warming in 1996, sea lamprey have been observed regularly, and the numbers recorded have increased in recent years. The scale shown in Figure 7 ends at 10, but in fact the numbers observed are higher because the registrations in some cases (e.g. few, many) do not allow exact numbers to be stated. Twaite shad were observed in 1994 and then annually between 2001 and 2004, but they have not been caught more recently. Ocean sunfish and garfish were recorded, respectively, in 1998 and 2000, although since then they have been observed irregularly.
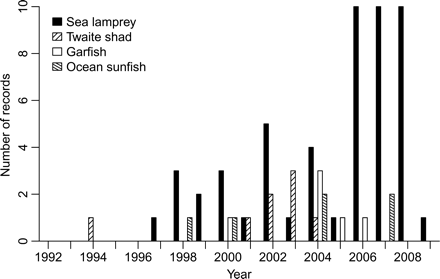
Records of some vagrant fish species in Icelandic waters during the period 1993–2010.
In addition to the species mentioned above, Ray's bream or Atlantic pomfret (Brama brama) is a vagrant species that has been caught in exceptional numbers recently. Ray's bream is a pelagic or mesopelagic species which in the eastern Atlantic is distributed from off the coast of southwestern Africa north to the British Isles. Occasionally, it has been found to the west of the British Isles, in the North Sea, and off the coast of Norway (Jonsson and Palsson, 2006). It has been found too in Icelandic waters several times but until recently had been absent for several decades. In late summer 2006, an Icelandic fishing boat caught ∼50 Ray's bream during a linefishery ∼740 m deep south of Iceland (Palsson, 2008). During summer 2008, Ray's bream was again caught along most of the south coast of Iceland at depths of 100–670 m and in various fishing gears (Palsson, 2009), and 34 individuals were brought to the Marine Research Institute for investigation. In 2009, Ray's bream was caught south of Iceland in exceptional numbers which, for the first time, resulted in a formally registered landed Icelandic catch of 3.5 t (Palsson, 2010). Before the above-mentioned catches, there is only one record from Icelandic waters when more than a single fish was caught—in 1953 (Jonsson and Palsson, 2006). This was at a time when warm climate conditions also prevailed in the North Atlantic Ocean.
Records of new species
During the recent warm period from 1996 to 2010, 31 fish species have been recorded from Icelandic waters for the first time. Table 1 lists these first recordings of new species in chronological order and also shows additional records for several of them during this period, along with information related to the catches. The table shows that one or more new species have been caught every year during this period, except for the years 2003, 2005, 2009, and 2010. In all, 17 species have been caught only as single individuals (Table 1). Eight species (black devil anglerfish, legless searsid, violet cuskeel, pink sabretooth, deepscale dory, palebelly searsid, Chaunax suttkusi, and Lycodes terraenovae) have been caught 2–9 times, but always as single individuals (Table 1). Only six species (blue shark, flounder, Poromitra megalops, sailfin dory, longspined bullhead, and four-spot megrim) have been caught as more than a single individual at a given station. The species that have been observed most often or in largest numbers are discussed below in more detail. Additional discussion on the other species is to be found in Jonsson and Palsson (2006) and Astthorsson and Palsson (2006).
Blue sharks were caught for the first time during summer 1996 on a line deployed by Japanese tuna boats conducting experimental tuna fishing within the Icelandic EEZ far south of Iceland (Astthorsson and Palsson, 2006). During the period 1997–2005, blue sharks were further found to be a frequent bycatch, and the number caught per season ranged from 30 to 846. The greatest abundance was at 60–62°N 16–24°W, and the northernmost records are from ∼50 miles off the south coast (Jonsson and Palsson, 2006). Most likely, this distribution reflects more the distribution of the effort in the tuna fishery than the actual distribution of blue sharks. This further seems to indicate that blue sharks are fairly common in the oceanic waters south of Iceland, and the recent record is probably mainly related to the new exploratory fishing rather than the warming climate.
Flounder were first caught with certainty near the mouth of the river Olfusa on the southwest coast of Iceland in September 1999 (Astthorsson and Palsson, 2006). Previously, during the same year, a suspicious “plaice/dab” had been caught by farmers in the vicinity, but was not brought to the Marine Research Institute for inspection. Since then, the flounder has been recorded almost annually and from many different locations all around the country, except for the northeast and east coasts (Jonsson et al., 2001). During spring 1997, spawning flounder were caught for the first time in a Danish seine in coastal waters off the west coast (Palsson, 2008). It is evident that the flounder has become distributed widely and rapidly in Icelandic coastal waters during the period since it was first recorded. This finding is of particular interest because flounder are mainly confined to shallow water and estuaries, and the records at Iceland are the westernmost for the species in the North Atlantic Ocean. Previously, the nearest records were from the Faroe Islands (Joensen and Tåning, 1970). The flounder is widely distributed along the coasts of mainland Europe, from the White Sea in the north to the coasts of Morocco in the south and is also found in the North Sea and the Baltic (Nielsen, 1986a).
In June 2000, ten P. megalops were caught at two stations in the Irminger Sea southwest of Iceland (Astthorsson and Palsson, 2006). Poromitra megalops is a mesopelagic species widely distributed in the northeastern North Atlantic Ocean south of Iceland, but these are to our knowledge the first confirmed records in Icelandic waters.
Sailfin dory were first caught in April 2001 in shallow water north of Reykjanes (Astthorsson and Palsson, 2006). In 2008, six more were caught at five stations off the south and west coasts (Palsson, 2009). Sailfin dory in the eastern Atlantic are distributed from the coasts of South Africa north to the west coast of Ireland (Quero et al., 1998; Swaby and Potts, 1999). During recent decades, sailfin dory have been found more frequently in northern European waters, and increasing sea temperature has been suggested as a factor in allowing the extended distribution (Swaby and Potts, 1999). A related species, John dory, also seems to be extending its distribution northwards in recent years. It was recorded for the first time in the waters southwest of Iceland in 2004, and in 2001, it was recorded in Faroese waters for the first time, and then again in 2006 (R. Mouritsen, pers. comm.).
Five longspined bullhead were caught in a littoral pool near Reykjavík in February and April 2006. The Marine Research Institute was informed about a similar fish from a nearby littoral area in 2005, but this record could not be confirmed. Longspined bullhead are distributed from the western Mediterranean north along the coasts of Europe and Scandinavia and into the North Sea and Baltic. They are also known from the Faroe Islands and now from Iceland (Fedorov, 1986; Palsson, 2007).
Six four-spotted megrim were caught at four stations at depths between 140 and 240 m on the shelf southwest of Iceland during April 2008 (Palsson, 2009). The fish ranged from 21 to 34 cm long, four were females, and two were males. All these fish were sexually mature and near spawning. In spring 2009, a further two were caught at a location near the 2008 catches. In the northeastern Atlantic Ocean, four-spot megrim are distributed from off the coast of Morocco in the south to Portugal, Spain (Bay of Biscay) and west of the British Isles (Nelson, 1986) in the north.
Concluding remarks
The results in the present study demonstrate that marked changes in distribution and abundance are currently taking place in the fish fauna around Iceland. These changes relate to exploited fish, less common species, and previously recorded vagrant species in Icelandic waters. Additionally, the first confirmed records of many new species in Icelandic waters have been made. Of the new records, we believe that only three (blue shark, carapine grenadier, and L. terraenovae) have been caught as a result of new exploratory fishing in deep water or because a new gear was used (Astthorsson and Palsson, 2006). In the cases of flounder and longspined bullhead, the possibility of transport by ballast water cannot be excluded. The other 26 species have not, despite continuous and heavy fishing for decades, been caught until now inside the Icelandic EEZ. Based on the geographic distribution, some of these 26 species could possibly be former inhabitants of the waters south and west of Iceland. However, similar to the changes observed for the exploited, less common, and vagrant species, we are of the opinion that many of the new records, and particularly those from heavily fished shelf areas (such as of Atlantic bonito, sailfin dory, John dory, and four-spotted megrim), reflect actual shifts that are taking place in the distribution of the fish fauna.
Most of the records of species that have never been observed before are from the Atlantic water off the south and west coasts of Iceland. Further, some of the new species are considered poor swimmers and are confined mostly to coastal or shelf waters, so the Icelandic records may demonstrate both the increased distribution of Atlantic water south and west of Iceland and the warming that has taken place during recent years.
Acknowledgements
We thank co-workers at MRI and the crews of RV “Bjarni Saemundsson” and RV “Arni Fridriksson”, who were involved in the hydrographic and fisheries surveys over the years. Also, we thank Icelandic fishers for collecting rare fish and bringing them to the Marine Research Institute for further investigation and Asta Gudmundsdottir for providing mackerel information.
References
Author notes
Handling editor: Bill Turrell

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}