





Abstract
Accumulating evidence shows that fishing exploitation and environmental variables can synergistically affect the population dynamics of exploited populations. Here, we document an interaction between fishing impact and climate variability that triggered a synchronic response in the population fluctuations of six exploited species in the Mediterranean from 1965–2008. Throughout this period, the fishing activity experienced a sharp increase in fishing effort, which caused all stocks to shift from an early period of underexploitation to a later period of overexploitation. This change altered the population resilience of the stocks and brought about an increase in the sensitivity of its dynamics to climate variability. Landings increased exponentially when underexploited but displayed an oscillatory behaviour once overexploited. Climatic indices, related to the Mediterranean mesoscale hydrography and large-scale north Atlantic climatic variability, seemed to affect the species with broader age structure and longer lifespan, while the global-scale El Niño Southern Oscillation index (ENSO) positively influenced the population abundances of species with a narrow age structure and short lifespan. The species affected by ENSO preferentially inhabit the continental shelf, suggesting that Mediterranean shelf ecosystems are sensitive to the hydroclimatic variability linked to global climate.Quetglas, A., Ordines, F., Hidalgo, M., Monserrat, S., Ruiz, S., Amores, Á., Moranta, J., and Massutí, E. 2013. Synchronous combined effects of fishing and climate within a demersal community. – ICES Journal of Marine Science, 70: 319–328.
Introduction
A current view among fisheries ecologists is that the effect of fishing exploitation on the population dynamics of harvested species cannot be separated from that of environmental variability (Perry et al., 2010; Planque et al., 2010). Interactions between fishing and climate trigger a variety of complex and often unpredictable biological responses. For instance, fishing exploitation may magnify the response of populations to environmental fluctuations by means of changes in demographic parameters, such as growth rate, which increase the instability of population dynamics (e.g. Hsieh et al., 2006; Anderson et al., 2008). In addition, demographic structure erosion makes populations more dependent on recruitment strength, and tightens the link between population and environmental variability (Botsford et al., 2011; Hidalgo et al., 2011; Rouyer et al., 2011). This evidence calls for a more comprehensive management approach that accounts for the combined effects of fishing and environment on ecological processes and population regulatory mechanisms.
Growing evidence from historical time-series analysis shows that climate can trigger large fluctuations in marine populations in the absence of fishing pressure, mainly described with respect to large and small pelagic species (Baumgartner et al., 1992; Ravier and Fromentin, 2001). However, the relationship between climate oscillations and fish populations has been demonstrated to be transitory, largely because of the non-stationary link between climate oscillations and regional-scale hydrography (Hsieh et al., 2009). In addition, natural populations fluctuate irrespective of the environmental scenario (i.e. relative contributions of low and high frequencies in the spectrum), because age-structure can act as a filter of environmental forcing (Bjornstad et al., 2004). Thus, depending on the species' life cycle and life history traits, recruitment signal spreads among age classes shaping the populations fluctuations (Bjornstad and Grenfell, 2001); ultimately, this results in a complex interplay between deterministic and stochastic processes. Most studies investigating how fishing affects this interplay focus on stock-specific case studies (but see CalCOFI database related studies, e.g. Hsieh et al., 2006; Anderson et al., 2008). However, fishing quite often harvests several species with a broad spectrum of life histories within a community, particularly the case for multi-specific fisheries, so a deep understanding of the way in which the same environmental variability can affect species with different life histories and demographic structure is required.
The Mediterranean is one of the most impacted upon seas in the world (Coll et al., 2008; Halpern et al., 2008; Lejeusne et al., 2010). Among ecological factors, such as direct habitat modification, species tropicalization, and seawater acidification (Calvo et al., 2011), fishing represents the activity having the greatest impact because it has driven most stocks to overexploitation (Colloca et al., 2011). Specifically, demersal fisheries, which are both multifleet and multispecific, demand an integrated multispecies approach to management (Moranta et al., 2008). In addition, marked cyclical fluctuations in the abundance of some species seem to be linked not only to exploitation, but also to the influence of climate on these populations (Oliver, 1993; Quetglas et al., 1998; Maynou, 2008b). Climate oscillations, such as the NAO (North Atlantic Oscillation), the MO (Mediterranean Oscillation) and IDEA (a recently described mesoscale index from the western Mediterranean) (Monserrat et al., 2008), have been reported to affect the cyclic abundance of demersal resources in the Mediterranean Sea (Lloret et al., 2001; Massutí et al., 2008; Maynou, 2008b).
In this study we aimed at investigating the combined effects of fishing and climate on the demersal community off the Balearic Islands (Western Mediterranean) using monthly information from 1965–2008 for six exploited species differing in life history characteristics and demographic structure. To do that, we first evaluated the temporal variation of the fishing exploitation level. Second, we investigated the oscillatory pattern of the time-series of population biomass between periods of different exploitation levels. Finally, we tested the potential influence of climatic variability on the population fluctuations.
Material and methods
Data sources
This study focuses on the bottom trawl fishery off Mallorca, the largest island of the Balearic Archipelago (western Mediterranean), that works on the continental shelf and upper slope, where it mainly targets striped red mullet Mullus surmuletus and European hake Merluccius merluccius, respectively. We analysed these two main target species and a set of four representative commercial by-catch species, for which long-term series of landings data were also available. Since the work is based on official statistics, the term “species” is used to refer both to the taxonomic species and to commercial categories. With the only exception being the category “elasmobranchs”, which includes the catshark, Scyliorhinus canicula, and the thornback ray, Raja clavata, the rest of the categories are composed chiefly of the following taxonomic species: red mullet, Mullus surmuletus; hake, Merluccius merluccius; octopus, Octopus vulgaris; squid, Loligo vulgaris; and cuttlefish, Sepia officinalis. Although we are aware of the limitations inherent in the use of commercial categories, we do not consider that they significantly affect the main conclusions of this work, because when there are mixtures of species in our study area, such mixtures contain sympatric or co-generic species having similar life-history traits. Consider, for instance, the worst case, the elasmobranch category composed of the catshark, S. canicula, and the skate, R. clavata. In our study area, both inhabit the same depth range (80–400m) (Massutí and Moranta, 2003) and share similar population characteristics (http://www.fishbase.org/): (i) maximum age: 9 years; (ii) trophic level: 3.69 vs 3.76; (iii) low resilience, with minimum population doubling time 4.5–14 years.
Monthly time-series of landings of these commercial species, along with the monthly fishing activity of each individual vessel from the trawling fleet of Mallorca during 1965 to 2008, were taken from official statistics. Communications with fishermen allowed us to use the actual, not nominal (declared), engine power (in HP) for each vessel, including changes in engine power over time and the corresponding increase in HP. The monthly fishing effort was obtained as the sum of the HP of all the vessels working during that month, weighted by the permitted time at sea (number of days per week and hours per day); the annual fishing effort (HP·y−1) was subsequently calculated by adding the monthly values. Previous to these calculations, however, we demonstrated that landings and effort acted proportionally during the time-series analysed, which is a required property if effort has to be summed for the entire fleet (Figure 1).
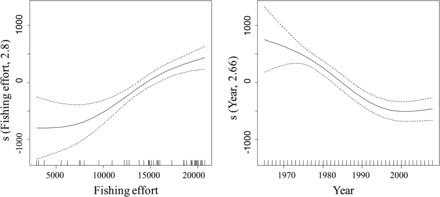
Results of a generalized additive model (GAM) describing landings as a function of fishing effort (in HP) and year. Landings and effort data correspond to the trawl fleet fishery off Mallorca (Balearic Islands, western Mediterranean) from 1965–2008. In the GAM, both effects (effort and year) were significant at p < 0.001, and the total deviance explained by the model was 71%.
In order to determine the influence of the climate on the population dynamics of the six stocks, we investigated three climatic indices of increasing spatial scale. The mesoscale (regional) IDEA index is a proxy of the hydroclimate in the northwestern Mediterranean (Monserrat et al. 2008). The IDEA index is based on the air/sea heat fluxes in the Gulf of Lion during the winter months, and was used as a proxy for the strength of the current in the northwest Mediterranean Sea (Monserrat et al., 2008). Lower values of the IDEA index indicate a larger formation of Western Mediterranean Intermediate Waters (WIW) in winter at intermediate layers (100–300 m depth) in the Gulf of Lion; this implies a larger presence of these water masses in the Balearic Sea during spring. The North Atlantic Oscillation (NAO) index, which is described as out-of-phase behaviour between the climatological low-pressure centre near Iceland and the high-pressure centre near the Azores, was used as a proxy of the North Atlantic climate (Hurrell, 1995). We used the winter average (December–March) of the NAO index (www.cru.uea.ac.uk/cru/data/nao.htm) because its influence in the Atlantic is higher during winter when the coupled ocean–atmosphere system is more active (Hurrell, 1995). The global-scale El Niño Southern Oscillation (ENSO) was also investigated by analysing the Southern Oscillation Index (www.cdc.noaa.gov/enso/enso.current.html), which measures the difference of sea level pressure between two localities situated in the eastern and central Pacific (Darwin and Tahiti, respectively). The NAO and ENSO indices have been demonstrated to influence the regional climate and the surface hydrographical conditions in the Mediterranean Sea (e.g. Mariotti et al., 2002; Hertig and Jacobeit, 2011).
Fishing exploitation
To analyse the temporal variation of the fishing exploitation level throughout the study period, a surplus production model was fitted to data (using the ASPIC software) for each selected species (Prager, 2004). In addition to the maximum sustainable yield (MSY), ASPIC estimates several benchmarks and stock status indicators such as population biomass (B), relative biomass (B/BMSY) and relative fishing mortality (F/FMSY); BMSY and FMSY stand for the B and F when stocks are exploited at their MSY. As a general consensus, the species with B/BMSY < 1 and F/FMSY > 1 are indicative of an overexploitation state, while B/BMSY > 1 and F/FMSY < 1 are indicative of an underexploitation state (Prager, 2004). ASPIC requires starting guesses of the following estimated parameters: (i) K, the stock's maximum biomass or carrying capacity; (ii) MSY, the maximum sustainable yield; (iii) B1/K, the ratio of the biomass at the beginning of the first year to K; and (iv) q, the catchability coefficient. In the first trials, we followed the recommendations of Prager (2004) in setting the starting guesses for these parameters, which were then interactively adjusted depending on the program outputs. The initial conditions for the MSY and K were half and ten times, respectively, the largest catch observed for each species during the period from 1965 to 2008. The starting value of the relative biomass (B1/K) was fixed at a range of values, as this can considerably reduce variance in the estimates and is a common practice when fitting surplus production models (Prager, 2004). The range of B1/K values used (0.5, 0.7, 0.9) reflects the lightly fished status of the stocks before 1965. For each estimated parameter, 80% bias-corrected confidence intervals were calculated using bootstrapping with 1000 iterations on the central set of estimates (B1/K = 0.7).
Population fluctuations
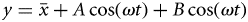
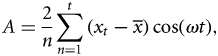
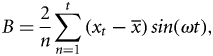
Climate effects
Climate effects on the population dynamics of the six stocks were investigated by means of Generalized Additive Models (GAMs). As a previous step to the GAM analyses, which bear the implicit assumption of additivity (covariates are mutually independent), we checked the lack of correlation among these three climatic indices (NAO vs IDEA: R = 0.099, p = 0.608; NAO vs ENSO: R = 0.229, p = 0.232; IDEA vs ENSO: R = 0.136, p = 0.482). To remove any possible effect of fishing effort in the oscillatory behaviour, the analyses were performed using CPUEs rather than landings. Furthermore, given that a decrease in the CPUEs series was observed for most species, which can be explained in part by increased fishing effort, the series were detrended (Maynou, 2008b). Detrending the original series renders this series stationary, and consisted of fitting an exponential regression to the data, with time as an independent variable (Abraham and Ledolter, 1983). The residuals obtained in these regressions were then used in the GAM analyses to model CPUEs as a function of the IDEA, NAO and ENSO indices. Previous to GAM analyses, Spearman correlations, accounting for temporal autocorrelations (Pyper and Peterman, 1998), were performed among species to inspect potential synchronic patterns.
Since in many cases climatic indices and CPUEs or landings are found to be correlated at different lag times (Lloret et al., 2001; Massutí et al., 2008; Maynou, 2008b), the GAMs were performed using lags from 0–7 years to include the highest periodicity reported for these indices (ENSO, 3–7 years). Consequently, eight (0–7 years) different GAMs were developed in which the abundance of each species was modelled against the IDEA, NAO, and ENSO indices as explanatory variables. GAM analyses were carried out with the R package (version 2.5.1; http://www.r-project.org/) using the mgcv library (Wood, 2001). The model selection criterion was based on minimizing the generalized cross validation (GCV), which measures the predictive squared error of the model; low values show the best compromise between model complexity and fit to the observed data (Wood, 2001). The goodness of fit in the GAMs was assessed using the percentage of deviance explained (DE) and the final GCV. For every model, residuals were checked for homogeneity of variance, absence of temporal autocorrelation and violation of normality assumptions.
Results
During the period from 1965–2008, the bottom trawl fishery off Mallorca showed large variations in the number of vessels, mean engine power and the fishing time at sea (Figure 2). Three main phases can be distinguished in the evolution of the total fleet fishing effort over time: (i) from 1965 to the mid 1970s it increased by a factor of 2.5; (ii) from the mid 1970s to 1994 it continued to grow but at a slower rate; and (iii) from 1994 to the present it has gradually decreased. Consistently for all species, time-series of landings showed a marked increase during the late 1970s that was followed by a progressive decrease, comparatively less pronounced for squid and cuttlefish (Figure 3). Although with some exceptions (occurring mainly during the first years of the time-series), annual CPUEs followed the same general pattern as the landings.
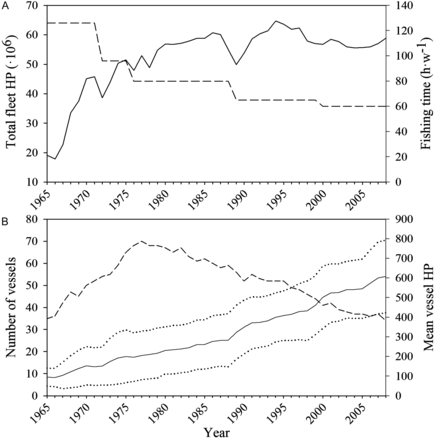
(A) Annual horse power (HP) (continuous line) of the entire bottom trawl fleet of Mallorca (Balearic Islands, western Mediterranean) and fishing time at sea (in hours per week, h·w−1) (stippled line) permitted by different regulations throughout the time-series 1965– 2008. (B) Total number of vessels (stippled line) along with mean and standard deviation (continuous and dotted lines, respectively) of mean vessel HP during the same time-series.
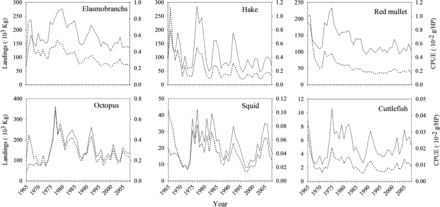
Landings (continuous line) and catch per unit effort (CPUE) (dotted line) of six demersal species caught by the bottom trawl fleet off Mallorca (Balearic Islands, western Mediterranean) during the period from 1965 to 2008.
Fishing exploitation
The evolution of the relative fishing mortality (F/FMSY) and biomass (B/BMSY) throughout the time-series showed a similar trend for all species (Figure 4). From the beginning of the series to the mid 1970s the stocks were underexploited, since the fishing effort was lower than the fishing effort that should be exerted to attain the maximum sustainable yield (F/FMSY < 1); consequently, the stocks' biomass was higher than the expected biomass at MSY (B/BMSY > 1). All the species turned to an overexploited state (i.e. B/BMSY < 1 and F/FMSY > 1) around the early 1980s, which continues to the present day. Although estimates of exploitation status of stocks expressed in relative terms were very similar for all the species studied (Supplementary Table S1), two main groups could be distinguished. The first group contained the two target species of the trawl fishery, red mullet and hake, whose central estimates of B2009/BMSY, F2008/FMSY, and Ye2009/MSY were 0.48, 1.4, and 0.73, respectively. The second group included the by-catch species (elasmobranchs, octopus, squid, and cuttlefish), whose central estimates for the same three parameters were 0.61–0.66, 1.08–1.58, and 0.85–0.89, respectively.
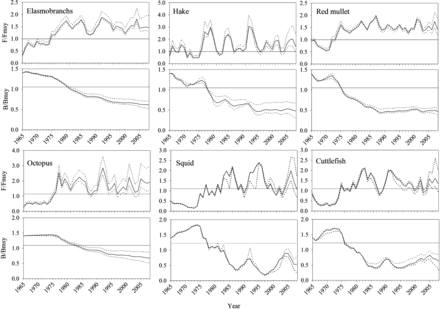
Time trajectories of the relative fishing mortality rate (F/FMSY) and relative population biomass (B/BMSY) estimated using non-equilibrium surplus production models for six different demersal species from Mallorca (Balearic Islands, western Mediterranean) under the assumption B1/K = 0.7. Dotted lines are 80% bias-corrected confidence intervals from bootstrapping.
Population fluctuations
The time-series analyses showed that analysing monthly landings or CPUEs rendered similar results. Both series showed an increase during the first period (1965–1980) with absence of interannual periodicities, best fitted in all species using a positive exponential function for landings and a negative exponential function for CPUEs. This period ended simultaneously for all species at the beginning of the 1980s, when an interannual oscillatory behaviour became apparent from 1980 to 2008. The best-fitting sinusoidal function for each species is superimposed on the 1980–2008 data series in Figure 5 which, for clarity purposes, only shows the landings fits. As previously mentioned, the most evident signal observed in the monthly series was the seasonal oscillation. This annual cycle, present throughout the entire series of all species, was clearly revealed by the Fourier analysis, and showed two main peaks at 12 and 6 months corresponding to the seasonal cycle and the first harmonic, respectively. The discrete Fourier transform and the harmonic analysis showed interannual oscillations of 7 years for four out of the six species analysed (elasmobranchs, hake, octopus, and cuttlefish). In the case of squid and red mullet these oscillations were longer (11 and 15 years, respectively).
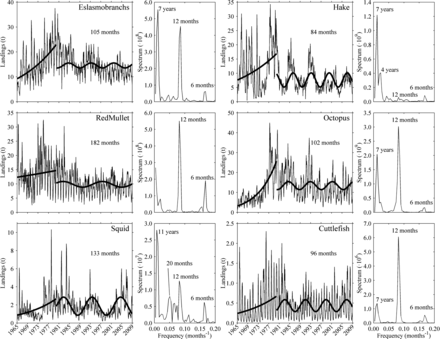
Spectrum analysis and least squares fitting to the monthly data (landings) of six demersal species caught by the bottom trawl fleet off Mallorca (Balearic Islands, western Mediterranean) during the period from 1965 to 2008. The best fit to the data is shown superimposed on the landings: an exponential function for the series 1965–1980, and a sinusoidal function for 1980–2008. The periodicity of the oscillations (in months) found in this last period is also shown for each species.
Climate effects
Residuals retained after removing the non-linear trend were significantly correlated in most species pairs (Table 1). Such residuals were then used in the GAM analyses to model CPUEs as a function of the IDEA, NAO and ENSO indices. To determine whether the fluctuations in the population dynamics were due to climate variability, we focused our analysis on the overexploited and oscillatory period (1980–2008). For each of the six species investigated, eight GAM models were performed, corresponding to the eight lag times (0–7 years) considered. Model selection and outputs of all these 48 models (8 lags × 6 species) are displayed in Supplementary Table S2, whereas Table 2 shows exclusively the best model for each species. Consistent results were obtained for four out of the six species (red mullet, octopus, squid, and cuttlefish), whose best model showed a significant positive effect of the ENSO index at a lag time of 6 years. The variance explained for these four models ranged from 26–33%. The two remaining species only showed significant effects of the NAO (hake at lag times of 2 and 5 years) and IDEA (hake and elasmobranchs at lag times of 7 and 2 years, respectively) indexes.
Correlation of residuals among the six species investigated (elasmobranchs, hake, red mullet, octopus, squid, and cuttlefish) obtained from fitting an exponential regression to CPUE data with time as an independent variable.
| Elasmobranch | Hake | Red mullet | Octopus | Cuttlefish | |
|---|---|---|---|---|---|
| Hake | 0.62*** | ||||
| Red mullet | 0.17 | –0.13 | |||
| Octopus | 0.61*** | 0.60*** | 0.14 | ||
| Cuttlefish | 0.39* | –0.02 | 0.30 | 0.61*** | |
| Squid | 0.50*** | 0.25 | 0.15 | 0.62*** | 0.57*** |
| Elasmobranch | Hake | Red mullet | Octopus | Cuttlefish | |
|---|---|---|---|---|---|
| Hake | 0.62*** | ||||
| Red mullet | 0.17 | –0.13 | |||
| Octopus | 0.61*** | 0.60*** | 0.14 | ||
| Cuttlefish | 0.39* | –0.02 | 0.30 | 0.61*** | |
| Squid | 0.50*** | 0.25 | 0.15 | 0.62*** | 0.57*** |
The residuals obtained in these regressions were then used in the GAM analyses to model CPUEs as a function of the IDEA, NAO and ENSO indices. *p < 0.05; **p < 0.005; p < 0.001.
Correlation of residuals among the six species investigated (elasmobranchs, hake, red mullet, octopus, squid, and cuttlefish) obtained from fitting an exponential regression to CPUE data with time as an independent variable.
| Elasmobranch | Hake | Red mullet | Octopus | Cuttlefish | |
|---|---|---|---|---|---|
| Hake | 0.62*** | ||||
| Red mullet | 0.17 | –0.13 | |||
| Octopus | 0.61*** | 0.60*** | 0.14 | ||
| Cuttlefish | 0.39* | –0.02 | 0.30 | 0.61*** | |
| Squid | 0.50*** | 0.25 | 0.15 | 0.62*** | 0.57*** |
| Elasmobranch | Hake | Red mullet | Octopus | Cuttlefish | |
|---|---|---|---|---|---|
| Hake | 0.62*** | ||||
| Red mullet | 0.17 | –0.13 | |||
| Octopus | 0.61*** | 0.60*** | 0.14 | ||
| Cuttlefish | 0.39* | –0.02 | 0.30 | 0.61*** | |
| Squid | 0.50*** | 0.25 | 0.15 | 0.62*** | 0.57*** |
The residuals obtained in these regressions were then used in the GAM analyses to model CPUEs as a function of the IDEA, NAO and ENSO indices. *p < 0.05; **p < 0.005; p < 0.001.
Results of generalized linear models (GAMs) fitting the abundance indices of six commercial species from the Mallorca trawling fleet (Balearic Islands, western Mediterranean) as a function of three different climatic indices with several lag times.
| Species | Lag time (years) | IDEA | NAO | ENSO | DE (%) |
|---|---|---|---|---|---|
| Red mullet | 6 | n.s. | n.s. | 0.024 | 26.0 |
| Octopus | 6 | n.s. | n.s. | 0.008 | 33.3 |
| Squid | 5 | n.s. | n.s. | 0.011 | 30.3 |
| 6 | n.s. | n.s. | 0.013 | 30.2 | |
| Cuttlefish | 0 | n.s. | n.s. | 0.052 | 16.8 |
| 6 | n.s. | n.s. | 0.010 | 31.9 | |
| Hake | 2 | n.s. | 0.022 | n.s. | 25.5 |
| 5 | n.s. | 0.065 | n.s. | 18.7 | |
| 7 | 0.030 | n.s. | n.s. | 38.5 | |
| Elasmobranch | 2 | 0.043 | n.s. | n.s. | 19.2 |
| Species | Lag time (years) | IDEA | NAO | ENSO | DE (%) |
|---|---|---|---|---|---|
| Red mullet | 6 | n.s. | n.s. | 0.024 | 26.0 |
| Octopus | 6 | n.s. | n.s. | 0.008 | 33.3 |
| Squid | 5 | n.s. | n.s. | 0.011 | 30.3 |
| 6 | n.s. | n.s. | 0.013 | 30.2 | |
| Cuttlefish | 0 | n.s. | n.s. | 0.052 | 16.8 |
| 6 | n.s. | n.s. | 0.010 | 31.9 | |
| Hake | 2 | n.s. | 0.022 | n.s. | 25.5 |
| 5 | n.s. | 0.065 | n.s. | 18.7 | |
| 7 | 0.030 | n.s. | n.s. | 38.5 | |
| Elasmobranch | 2 | 0.043 | n.s. | n.s. | 19.2 |
*p < 0.05, **p < 0.005, ***p < 0.001, n.s. = non-significant.
The following indices of increasing spatial scale were used: the IDEA index, the North Atlantic Oscillation (NAO) and the El Niño/Southern Oscillation (ENSO). The significance of each index and the best model fit based on the percentage of deviance explained (DE) are shown. The GAMs displayed were performed on the statistically significant fittings resulting from the entire set of 48 models considering eight lag times (0 to 7 years) (see Supplementary Table S2).
Results of generalized linear models (GAMs) fitting the abundance indices of six commercial species from the Mallorca trawling fleet (Balearic Islands, western Mediterranean) as a function of three different climatic indices with several lag times.
| Species | Lag time (years) | IDEA | NAO | ENSO | DE (%) |
|---|---|---|---|---|---|
| Red mullet | 6 | n.s. | n.s. | 0.024 | 26.0 |
| Octopus | 6 | n.s. | n.s. | 0.008 | 33.3 |
| Squid | 5 | n.s. | n.s. | 0.011 | 30.3 |
| 6 | n.s. | n.s. | 0.013 | 30.2 | |
| Cuttlefish | 0 | n.s. | n.s. | 0.052 | 16.8 |
| 6 | n.s. | n.s. | 0.010 | 31.9 | |
| Hake | 2 | n.s. | 0.022 | n.s. | 25.5 |
| 5 | n.s. | 0.065 | n.s. | 18.7 | |
| 7 | 0.030 | n.s. | n.s. | 38.5 | |
| Elasmobranch | 2 | 0.043 | n.s. | n.s. | 19.2 |
| Species | Lag time (years) | IDEA | NAO | ENSO | DE (%) |
|---|---|---|---|---|---|
| Red mullet | 6 | n.s. | n.s. | 0.024 | 26.0 |
| Octopus | 6 | n.s. | n.s. | 0.008 | 33.3 |
| Squid | 5 | n.s. | n.s. | 0.011 | 30.3 |
| 6 | n.s. | n.s. | 0.013 | 30.2 | |
| Cuttlefish | 0 | n.s. | n.s. | 0.052 | 16.8 |
| 6 | n.s. | n.s. | 0.010 | 31.9 | |
| Hake | 2 | n.s. | 0.022 | n.s. | 25.5 |
| 5 | n.s. | 0.065 | n.s. | 18.7 | |
| 7 | 0.030 | n.s. | n.s. | 38.5 | |
| Elasmobranch | 2 | 0.043 | n.s. | n.s. | 19.2 |
*p < 0.05, **p < 0.005, ***p < 0.001, n.s. = non-significant.
The following indices of increasing spatial scale were used: the IDEA index, the North Atlantic Oscillation (NAO) and the El Niño/Southern Oscillation (ENSO). The significance of each index and the best model fit based on the percentage of deviance explained (DE) are shown. The GAMs displayed were performed on the statistically significant fittings resulting from the entire set of 48 models considering eight lag times (0 to 7 years) (see Supplementary Table S2).
Discussion
This study showed that the population dynamics of the main commercial species of the demersal community off the Balearic Islands has been synchronously affected by the combined effect of fishing exploitation and environment variability during the last forty-four years. Such a synchronic effect has been twofold. First, the fact that all the species here studied became overexploited (i.e. B/BMSY < 1) around the early 1980s seems unlikely to be an exclusive effect of fishing. We hereafter discuss how changes in the Mediterranean hydroclimate in the early 1980s could have contributed to synchronizing the changes observed. And second, by turning to an overexploited state, populations seem to have been more susceptible to environmental variability (Perry et al., 2010; Planque et al., 2010), displaying synchronic fluctuations in the population biomass.
The six species analysed followed a general trend characterized by an early period of underexploitation that shifted to overexploitation in the early 1980s. This community shift affected the population dynamics of all the species, since their landings and CPUEs showed an increasing trend during the underexploitation phase (1965–1980) but an oscillatory behaviour during the overexploitation phase (1980–2008). These natural fluctuations, however, would have been masked during the first part of the series, when landings displayed an exponential growth due to the rapid increase in fishing effort. Therefore, our results indicate a phase-transition event in the population dynamics of demersal stocks from the Balearic Islands in 1980, a scenario already reported for hake in this area (Hidalgo et al., 2009). The rapid increase in the exploitation rate observed during the previous years (namely, it increased 2.5 times during 1965–1977) seems to have had the major effect of sudden depletion. However, growing evidence shows that the hydroclimate of the northwest Mediterranean also changed suddenly, around the early 1980s, to an overall dominance of higher temperatures and atmospheric pressure, and weaker wind stress and precipitation (Fernandez de Puelles and Molinero, 2007, 2008; Molinero et al., 2009; Conversi et al., 2010). These changes in the hydrography influenced the seasonal and interannual plankton dynamics around the Balearic Islands (Fernandez de Puelles and Molinero, 2007, 2008) that could have affected the survival of early life stages of exploited species, and, in turn, synergistically magnified the effect of fishing (Hidalgo et al., 2011).
Fluctuations in landings of demersal species in the western Mediterranean are well documented. Astudillo and Caddy (1986) found a marked periodicity of 12 years in the landings of hake and red mullet from the Balearic Islands, which they hypothesized to be independent of fleet activity. In the same area, a periodicity of about 8 years was reported for octopus (Quetglas et al., 1998) and red shrimp (Carbonell et al., 1999). In a nearby area, however, fluctuations in red shrimp landings ranged between 7 and 13 years among different ports (Maynou, 2008a). Our results showed interannual oscillations with a periodicity of 7–9 years in the last three decades for four out of the six species investigated. There is increasing evidence that harvested species fluctuate more than unharvested ones, which is probably due to the elevated variability of recruitment and the increased sensitivity to environmental variability resulting from the demographic truncation caused by fishery exploitation (Hsieh et al., 2006; Anderson et al., 2008).
To determine whether the causes of the interannual fluctuations in CPUEs were linked to climatic variability, climate indices at increasing spatial scales (local, meso-, and global-scale) were examined. NAO and IDEA showed significant effects on the elasmobranches and hake consistent with previous studies (Massutí et al., 2008). Unexpectedly, however, our models found much support for an effect of ENSO with a lag time of 6 years for four out of the six species analysed (red mullet, octopus, squid, and cuttlefish). Identical responses in these four species (Figure 5) point to an ENSO signal in some Mediterranean marine populations. This signal might, in fact, interact with, or have masking effects on, other influences such as the NAO, which has displayed a multidecadal component of ca. 8 years during recent decades (e.g. Massei et al., 2007 and references therein), close to the periodicity found in most of our time-series (7 years). Indeed, the interaction between North Atlantic climate (i.e. NAO) and Pacific modes (i.e. ENSO) has been shown to affect the Mediterranean climate through rainfall (e.g. Mariotti et al., 2002). Recent evidence also demonstrated the influence of ENSO variability on Mediterranean hydrographical characteristics, particularly on the sea surface temperature (Hertig and Jacobeit, 2011). In the Balearic Islands, rainfall and sea surface temperature variability strongly affect the nutrients, phytoplankton, and zooplankton variability (Fernandez de Puelles and Molinero, 2007, 2008). Since the species displaying a close link with ENSO are those inhabiting the shallow continental shelf, they should be more influenced by the interannual dynamics of zooplankton affecting prey-predator relationships and survival of early life stages. Links between ENSO and coastal species outside the Mediterranean are not new and have been reported for other organisms worldwide (Urban et al., 2000; Rimbu et al., 2003).
Though a time lag between the climatic and the biological signal was expected (e.g. Ottersen et al., 2004; Overland et al., 2010), the 6-year lag of the ENSO effect on the population biomass of the four continental shelf species is a striking result of our study (but see, Gonzalez-Herraiz et al., 2009; Zimmerman and Palo, 2012). Since the ENSO signal on the Mediterranean hydroclimate is transmitted within some months (Hertig and Jacobeit, 2011), there must be additional factors modulating the population fluctuations that we have not taken into account in our study. We suggest that internal properties of the life cycle of these species can interact with the climate signal, which makes plausible potential inter-cohort and density-dependent effects (Bjornstad and Grenfell, 2001). Favourable climatic events can produce strong annual cohorts, favouring the recruitment in consecutive years due to the relative good level of reproductive stock. This could magnify the climatic signal in the next generations and thus bring forth population cycles (Tzeng et al., 2012) that could explain the lagged oscillatory response observed.
Recent studies on species with a broad demographic structure have shown that variability in recruitment increases with fishing and becomes particularly sensitive to forcing at time-scales near the mean age of reproduction (Botsford et al., 2011). This is consistent with the cohort resonance effect (Bjornstad et al., 1999, 2004) and has been recently suggested for hake in the Balearic Islands (Hidalgo et al., 2011). Our study adds to this picture of how species of the same harvested community with different life history and demographic structure cope with environmental variability. Our results support the hypothesis that population fluctuations result from the interaction of deterministic and stochastic processes (Bjornstad and Grenfell, 2001), while the relative contribution of each component depends on the life history and demographic structure of the species. Hake, red mullet, and elasmobranch species have a broader age structure and longer life span compared with squid, cuttlefish, and octopus, and thus deterministic mechanisms (e.g. internal demographic processes) may have higher impact on population fluctuations. By contrast, cephalopods display a narrow age structure, fast life cycle and short life span, and thus population fluctuations are likely to be more affected by climate variability. It is worth noting that our results suggest potential effects of the same climatic phenomena at different temporal lags that may be further evaluated in a unique model structure for each species, as well as potential interactions among the different climatic indices. Future studies should tackle this challenge, combining long-term CPUE data with age-structure (stage-structure) information for assessment groups that will allow investigation as to whether and how climate can affect the same species through several demographic processes.
To conclude, our results demonstrate that interaction between fishing and climate can have synchronic effects on the population fluctuations of different species within an overexploited community. Intense fishing exploitation may increase the susceptibility, and therefore reduce the resilience, of harvested populations to climate variability (Botsford et al., 2011). We have also shown that, as a consequence of this increased susceptibility, populations may increase their sensitivity to subtle climate signals that could otherwise have been masked by more influential factors. Our study expands this conclusion to the community scale, and indicates that sensitivity to climate, and thus population fluctuations, depend on the demographic characteristics, life history, and bathymetric distribution of each species.
Supplementary Data
Supplementary material is available at the ICESJMS online version of the paper:
Supplementary Table S1. Surplus-production model results.
Supplementary Table S2. Outputs of 48 GAMs (8 lags × 6 species) modelling CPUEs as a function of the climatic indexes IDEA, NAO, and ENSO.
Funding
This study was financed by the PROGECIB-48A project, funded by the Direcció General de R + D + I (Conselleria d' Economia, Hisenda i Innovació, Govern de les Illes Balears).
Acknowledgements
We would like to express our gratitude to all the Confraries de Pescadors de Mallorca, the Instituto Social de la Marina and the Federació de Confraries de Mallorca, for kindly providing data on landings and fleet activity from the different ports of the island. Thanks to M. H. Prager (NOAA, North Carolina, USA) for his help with the ASPIC software. Special thanks to Pep Domingo, retired fisherman from Mallorca, for giving us information about fleet activity, vessel characteristics, and life on board in the fifties, when he began to fish. The English version was revised by C. Stonehouse.
References
Author notes
Handling editor: Howard Browman

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}