





Abstract
Testing models of macroevolution, and especially the sufficiency of microevolutionary processes, requires good collaboration between molecular biologists and paleontologists. We report such a test for events around the Late Cretaceous by describing the earliest penguin fossils, analyzing complete mitochondrial genomes from an albatross, a petrel, and a loon, and describe the gradual decline of pterosaurs at the same time modern birds radiate. The penguin fossils comprise four naturally associated skeletons from the New Zealand Waipara Greensand, a Paleocene (early Tertiary) formation just above a well-known Cretaceous/Tertiary boundary site. The fossils, in a new genus (Waimanu), provide a lower estimate of 61–62 Ma for the divergence between penguins and other birds and thus establish a reliable calibration point for avian evolution. Combining fossil calibration points, DNA sequences, maximum likelihood, and Bayesian analysis, the penguin calibrations imply a radiation of modern (crown group) birds in the Late Cretaceous. This includes a conservative estimate that modern sea and shorebird lineages diverged at least by the Late Cretaceous about 74 ± 3 Ma (Campanian). It is clear that modern birds from at least the latest Cretaceous lived at the same time as archaic birds including Hesperornis, Ichthyornis, and the diverse Enantiornithiformes. Pterosaurs, which also coexisted with early crown birds, show notable changes through the Late Cretaceous. There was a decrease in taxonomic diversity, and small- to medium-sized species disappeared well before the end of the Cretaceous. A simple reading of the fossil record might suggest competitive interactions with birds, but much more needs to be understood about pterosaur life histories. Additional fossils and molecular data are still required to help understand the role of biotic interactions in the evolution of Late Cretaceous birds and thus to test that the mechanisms of microevolution are sufficient to explain macroevolution.
Introduction
The question whether microevolutionary processes (that can be studied in the present) are sufficient to account for all of macroevolution is still debated (Simons 2002) and, in particular, whether mammals and birds would have replaced dinosaurs and mammals even without the extraterrestrial impact that marks the K/T (Cretaceous/Tertiary) boundary (see Penny and Phillips 2004). Patterns of diversity and processes of ecological partitioning in Late Cretaceous birds and flying reptiles are incompletely known. It is clear that early crown birds (Neornithes) overlapped with archaic birds (such as the enantiornithes and the diving birds Hesperornis and Ichthyornis) as well as with pterosaurs. Furthermore, none of the latter groups—except for the crown birds—survived into the Tertiary. There was a decline to extinction of both pterosaurs and archaic birds such as Hesperornis and Ichthyornis, but did this precede or follow the rise of modern (crown group) birds (Neornithes)? In order to test models (Penny and Phillips 2004) about extinctions around the end of the Cretaceous we need to understand important evolutionary events before the asteroid impact that marks the Cretaceous/Tertiary boundary (Cooper and Penny 1997; Cracraft 2001; Van Tuinen and Hedges 2001; Wilf et al. 2003; Peters 2005). Hypotheses for the origin of modern (crown group) birds vary considerably (Penny and Phillips 2004). Did just one lineage survive the K/T boundary extinctions (Feduccia 2003) and then diversify rapidly, or did many modern lineages diversify from early in the Late Cretaceous, 80–100 Ma (Hedges et al. 1996; Cooper and Penny 1997; Härlid et al. 1997; Cracraft 2001; Paton et al. 2002; Dyke and van Tuinen 2004; Harrison et al. 2004)? To test these hypotheses we require additional well-defined calibration points, and penguins, with their excellent fossil record (Simpson 1975; Fordyce and Jones 1990; Myrcha et al. 2002; Clarke et al. 2003) are important. These large, aquatic, wing-propelled diving birds have solid (non-pneumatic) bones that preserve well, providing a sound fossil record of their lineage and dating their minimum divergence time from related flying birds.
Four associated fossil skeletons representing two new species of Early Paleocene penguins were found near Waipara (North Canterbury, New Zealand) (Fordyce and Jones 1990; Jones and Mannering 1997). They are from the Waipara Greensand, a well-dated unit from a thoroughly described sequence (Vajda et al. 2001; Hollis and Strong 2003) that includes an important K/T site. Indeed it is near one of the three classic K/T sites where the iridium anomaly was first reported (Alvarez et al. 1980). The older of the two new species is the oldest fossil penguin found thus far, and is therefore important for both understanding the origin and evolution of penguins (Baker et al. 2006), as well as providing an improved calibration point for estimating the early divergence times of modern sea and shorebirds. The late Early Paleocene to Late Paleocene age of these new fossils is important because it mitigates against any possible effects from any (hypothetical) coordinated acceleration of mutation rates of DNA sequences that might affect molecular estimates of divergence times (Conway Morris 1998; Foote et al. 1999; Bromham and Hendy 2000).
Given these excellent early penguin fossils, it is important to establish an evolutionary tree for penguins and their close relatives. We have reported a complete mitochondrial genome from the little blue penguin (Eudyptula minor) (Slack et al. 2003) but that study had a restricted number of taxa. Unexpectedly perhaps, though not inconsistent (on the present data set) with some earlier work (Sibley and Ahlquist 1990), storks were the sister taxon of penguins. Previously (Cooper and Penny 1997) we found that procellariforms (such as albatrosses, petrels, and shearwaters) were closer to penguins, but they were not represented at that time by complete mitochondrial genomes. We therefore sequenced the mitochondrial genomes of a small albatross (black-browed mollymawk, Diomedea melanophris), Kerguelen petrel (Pterodroma brevirostris), and red-throated loon (Gavia stellata). Some group other than procellariforms could still be the sister group to penguins (Cracraft 2001), but that does not affect the minimum age for the divergence of penguins from related lineages—our estimate is conservative in this respect. By combining the fossil and molecular data we report an excellent calibration point from penguin fossils, and use it to study questions regarding the early evolution of modern birds. To help understand some of the biotic interactions in the Late Cretaceous, we have considered the diversity and lifestyles of birds and of pterosaurs through the Cretaceous. The basic information for birds is from Chiappe and Dyke (2002) and Fountaine et al. (2005), and for pterosaurs from Wellnhofer (1991), supplemented with more-recent publications listed in Supplementary Material.
Our approach is in three parts: (1) we describe and analyze some excellent new penguin fossils, (2) we report and analyze three new mitochondrial genomes of penguin relatives, and (3) we analyze some patterns and processes amongst pterosaurs to help elucidate events prior to the K/T boundary.
Description of the Oldest Fossil Penguins, Genus Waimanu
The fossil material represents two species of wing-propelled diving bird that are named and described here and illustrated in Figure 1. Each was found as a natural association of bones from a single individual, and judging from fusion of elements (vertebrae in synsacra) and some well-developed articular surfaces, they each represent a mature individual. The larger and older species is almost the size of an Emperor penguin (Aptenodytes forsteri); the smaller and geologically slightly younger is estimated at 80 cm tall, about the size of a yellow-eyed penguin (Megadyptes antipodes). Most parts of the skeleton are preserved in Waimanu (fig. 1), allowing ready comparison with literature on avian cladistics (Cracraft 1988; Mayr and Clarke 2003). Cladistic analyses, given below, place Waimanu at the base of the Sphenisciformes (sensu Clarke et al. 2003); and identify characters used in the generic diagnosis below.
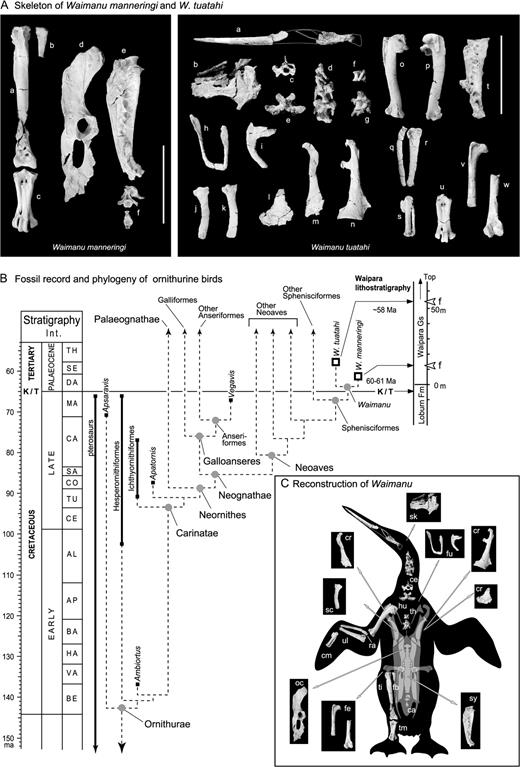
The Paleocene penguin Waimanu. (A) Skeletal elements of Waimanu. Left box, W. manneringi. a. right tibiotarsus in anterior view, b. right fibula in anterior view, c. right tarsometatarsus in dorsal view, d. right os coxae in lateral view, e. synsacrum in lateral view, f. caudal vertebrae in anterior view (a–f are CM zfa 35). Right box, W. tuatahi. a. mandible in lateral view (anterior part) and in medial view (posterior part), b. cranium in dorsal view, c–e. cervical vertebrae in anterior view (c), and in ventral view (d, e), f–g. thoracic vertebrae in lateral view, h. furcula in anterior view, i. right/left clavicle in lateral view, j–k. scapulae in lateral view, l. right coracoid in ventral view, m–n. left coracoids in ventral view, o. right humerus in ventral view, p. right humerus in dorsal view. q. left radius in dorsal view, r. left ulna in dorsal view, s. left carpometacarpus in dorsal view, t. synsacrum in lateral view, u. right tarsometatarsus in dorsal view, v. right femur in anterior view, w. left femur in posterior view. (a, c, h, j, l–m, p–r, t, and v are OU 12651; f–g, i, k, n, o, s, u, and w are CM zfa 34; b, d, and e are zfa 33). Scale bar = 100 mm. (B) Fossil record and phylogeny of ornithurine birds with the stratigraphy of Waipara region and geological settings for Waimanu. Solid line shows geological ranges of taxa with first and last occurrences shown by squares. Dashed line shows postulated phylogeny compiled from literature (Martin and Stewart 1982; Fox 1984; Chiappe 1995, 2003; Elzanowski et al. 2000; Norell and Clarke 2001; Cracraft and Clark 2001; Chiappe and Dyke 2002; Clarke and Norell 2002; Galton and Martin 2003; Clarke 2004). Gray circles indicate possible initial divergence times for clades; known fossils (squares) show constraints on ages. Early divergences within the Carinatae could be older, and we have conservatively placed them later in the Cretaceous to give only one long ghost-lineage between Ambiortus and the early Carinatae. The placement of Waimanu within Sphenisciformes is evaluated by the cladistic analysis described in the text; see also supplementary figure 4. TH, Thanetian; SE, Selandian; DA, Danian; MA, Maastrichtian; CA, Campanian, SA; Santonian, CO, Coniacian; TU, Turonian; CE, Cenomanian; AL, Albian; AP, Aptian; BA, Barremian; HA, Hauterivian; VA, Valanginian; BE, Berriasian. (C) Reconstruction of Waimanu (composite of W. manneringi and W. tuatahi, based on original art by Chris Gaskin ©Geology Museum, University of Otago). ca, caudal vertebrae; ce, cervical vertebrae; cm, carpometacarpus; cr, coracoid; fb, fibula; fe, femur; fu, furcula; hu, humerus; sk, skull, md, mandible; oc, os coaxae; ra, radius; sc, scapula; sy, synsacrum; ti, tibiotarsus; tm, tarsometartarsus; ul, ulna. In the wing, the dorsal view (left ulna, radius, carpometacarpus) and ventral view (humerus) are combined.
Aves Linnaeus 1758 Neornithes Gadow 1893 sensu Cracraft 1988 Sphenisciformes Sharpe 1891 (sensu Clarke et al., 2003) Waimanu Jones, Ando and Fordyce, gen. nov.
Type species. Waimanu manneringi sp. nov.
Etymology. Maori: wai (water), manu (bird).
Included species. Type species and Waimanu tuatahi sp. nov.
Diagnosis. Waimanu is closer to penguins than to any other bird group in the following combination of characters: some thoracic vertebrae are not heterocoelous; synsacrum has 11–12 ankylozed vertebrae; hypotarsal crests and grooves of the tarsometatarsus are not well developed (but medial hypotarsal crest is distinct). Waimanu differs from other stem- and crown-Sphenisciformes in that: humerus has an elongated and elevated insertion of supracoracoideus, and scapulotricipital groove does not form a trochlea; ulna has ridged anterior proximal margin; radius lacks anterior angulation and notch; carpometacarpus has stepped anterior margin; scapular blade is evenly wide; thoracic vertebrae are very weakly opisthocoelous and laterally excavated; synsacrum has column-like vertebral bodies; femur has a deep patellar groove; extensor groove of tibiotarsus runs on medial side; and tarsometatarsus is long and waisted, with posterior-directed medial trochlea, distinct medial hypotarsal crest, and very shallow intermetatarsal grooves dorsally.
Waimanu manneringi Jones, Ando and Fordyce sp. nov.
Etymology. Honoring Al Mannering who found and collected the holotype.
Holotype. CM (Canterbury Museum) zfa35: associated part skeleton comprising almost complete right tibiotarsus, proximal half of right fibula, right tarsometatarsus, right pelvis, and synsacrum (with last thoracic vertebra attached to the synsacrum), four caudal vertebrae (fig. 1A).
Horizon, Locality, and Age. Basal Waipara Greensand (Wilson 1963; Field and Brown 1989), Waipara River (near 43°04′S, 172°36′E), New Zealand (Fossil Record Number M34/f453, NZ Fossil Record File). Calcareous nannofossils indicate late early Paleocene, 60.5–61.6 Ma (correlations after Cooper 2004); see Supplementary Material.
Diagnosis. Larger than W. tuatahi, with tarsometatarsus 78 mm long (cf. 65 mm).
Summary description. Tarsometatarsus: The tarsometatarsus is longer and more waisted than in more-crownward Sphenisciformes, but is still short and robust compared to volant birds. The prominent intercotylar projection is pointed proximally and dorsally. The proximal vascular foramina are small and, like the distal foramen, open onto the plantar surface. The hypotarsal crests are simplified as in other Sphenisciformes, and not enclosed to form a canal, though the medial crest is more pronounced than in other Sphenisciformes. Trochlea III is the longest, the other trochleae are slightly shorter and of subequal length, and trochlea IV is directed somewhat plantarly. The intermetatarsal grooves are separate, as in other Sphenisciformes, but very shallow.
Tibiotarsus: The tibiotarsus is broken in its distal third, revealing dense internal bone, but retains almost its full length; length is intermediate between King penguin (Aptenodytes patagonicus) and Emperor penguin (Aptenodytes forsteri). Profiles indicate that the cnemial crest protruded proximally. The shaft is more robust than in extant penguins, and the structure resembles that of Paraptenodytes antarcticus.
Os coxae: On the relatively complete right os coxae, the ilioischiadic foramen is larger than the acetabulum, and is closed posteriorly. The lateral margin of the preacetabular wing of the ilium has a deep concavity while the dorsal margin is rather straight. The dorsal iliac crest and the spinal crest of the synsacrum are not fused. The dorsolateral crest begins more posteriorly than in extant penguins. The preacetabular process is not well developed; the medial surface of the postacetabular part is rather flat, without a distinct renal fossa. Details are lost for the anterior margin of the ilium, the posterior and ventral margins of the ischium, and the pubis.
Synsacrum and caudal vertebrae: 11 fused vertebrae form this element; it is not fused with the os coxae. The spinal crest is high, robust, and full length. The vertebral bodies do not form a flat ventral surface at the middle of the synsacrum but keep a columnar structure. The synsacrum is intermediate in size between that of King penguin and Emperor penguin. The caudal vertebrae are unsurprising; one has a strong haemal process.
Waimanu tuatahi Ando, Jones and Fordyce sp. nov.
Etymology. Maori: tuatahi (first)—the holotype was the first specimen found.
Holotype. OU 12651 [Geology Museum, University of Otago]: associated part skeleton including skull fragments, incomplete mandible, cervical vertebrae, ribs, synsacrum, a furcula, coracoids, anterior half of right scapula, right humerus, distal end of left humerus, an ulna, a radius; proximal end of right femur (fig. 1A).
Hypodigm material. Holotype (OU 12651), and other associated part skeletons of: CM zfa 34—skull fragments, cervical and thoracic vertebrae, furcula, scapulae, coracoids, ribs, humeri, a radius, a carpometacarpus, and a tarsometatarsus (fig. 1A); CM zfa 33—partial skull and mandible, cervical and thoracic vertebrae, synsacrum, furcula, scapulae, coracoids, humerus, and a femur (fig. 1A).
Horizon and locality. All from the middle to upper Waipara Greensand, Waipara River. OU 12651, near 43° 02′ S, 172° 32′ E, fossil record number M34/f138; CM zfa 33, M34/f454; CM zfa 34, M34/f455 (zfa 33 and zfa 34, about 30 m stratigraphically above zfa 35; all zfa specimens are from near 43° 04′ S, 172° 36′ E). Dinoflagellates from M34/f138 indicate early Late Paleocene, 58–60 Ma (Cooper 2004: Fig. 11.5); M34/f454 and M34/f455 have not produced age-diagnostic fossils but are older than Early Eocene, no older than Paleocene, and stratigraphic level in upper Waipara Greensand indicates Late Paleocene; see Supplementary Material.
Specific diagnosis. Smaller than W. manneringi, with tarsometatarsus 65 mm long (cf. 78 mm long). On the tarsometatarsus, concavities of medial and lateral margins are less distinct, and plantar deflection of trochlea of metatarsal II is relatively weak.
Summary description. Skull: Where elements can be compared, there are no significant differences between OU 12651, zfa 34 and zfa 33. There are bilateral nasal gland fossae on the dorsal surface of the frontal. Frontal and parietal are fused completely, and the temporal fossa is large, reaching the top of the skull. The lacrimal has a descending process. The preserved part of the jugal bar is not dorsoventrally curved. On the quadrate, the otic process bears the separated articular surface for the otic and the squamosal. The palatine is present but incomplete.
Mandible: Partial mandibles are preserved. The symphysis is long and completely ossified. The anterior part of each ramus is slender and straight; the rami diverge only slightly posteriorly, but not enough is preserved to judge profiles toward the articulation. The medial mandibular fossa is large.
Furcula: The acromial process is well developed, and makes an acute angle with the rest of the clavicle. The articular facet of the acrocoracoid is projecting. The furcula lacks an interclavicular process, and probably did not abut the carina of the sternum.
Scapula: The acromion and glenoid process are rather symmetrical, not pronounced. The body is evenly wide posteriorly.
Coracoid: The shoulder end is elongate, with a medially bending acrocoracoid process; the scapular cotyla is a round shallow depression. The procoracoid process strongly projects medially, contributing to a thin and distally extended plate-like margin. Profiles indicate that a coracoid foramen was present. The sternal end is flared with a well-developed medial angle, and the base of a lateral process.
Wing elements: Overall, the wing is short relative to body size for birds in general, although relatively longer than in more-crownward penguins. Elements are variably flattened dorsoventrally, more so for the humerus than other bones. Broken sections reveal dense internal bone. The humerus has a sigmoidal shaft, and a well-developed articular surface excavated by a large and deep pneumotricipital fossa which lacks a pneumatic foramen at the base. The insertion of M. supracoracoideus is elevated and elongate distally, while a slight impression marks the origin for M. brachialis at the distal end of the anterior margin. The dorsal and ventral condyles are relatively prominent, and not flattened. The scapulotricipital and humerotricipital grooves are well developed, but only the latter forms a trochlea with strong ridges on the posterior margin of the distal end. The ulna is straight and broad proximally, while the radius is narrow and curved; these bones are shorter than the humerus (ca. 80% of length). The carpometacarpus is relatively broad and straight, with a stepped anterior margin.
Synsacrum: The synsacrum is smaller than but otherwise similar to that of W. manneringi.
Femur: The femur is long and straight, with only a slight posterior bend distally. The trochanteric crest is only weakly projected. The patellar groove is deep with distinct patellar crests. Broken sections reveal dense internal bone.
Tarsometatarsus: A nearly complete right tarsometatarsus (CM zfa 34), damaged proximally, is less waisted than in W. manneringi (namely, concavities of medial and lateral margins are less distinct) and is smaller (83% as long). Further, the trochlea of metatarsal II is relatively weakly deflected plantarly.
Remarks
Superficially, Waimanu is similar to geologically younger wing-propelled diving birds, such as the extinct Northern Hemisphere auk Mancalla and the diving plotopterid pelican Copepteryx (see Miller and Howard 1949; Olson 1980). Cladistically, Waimanu is closely related to modern penguins and thus belongs in the stem-Sphenisciformes (see below); they are large robust birds (ca. 80–100 cm), with dense heavy bones. Compared with volant birds, and as in other wing-propelled divers, the wing is short relative to body size, with flattened and generally wide bones; however, the distal ulna and radius are not widened. The structure of the humerus suggests limited rotation of the elbow; wing form rules out aerial flight and is consistent with wing-propelled diving. The structure of the humerus head and coracoid are reminiscent of other wing-propelled divers (Miller and Howard 1949; Howard 1966, 1970, 1976; Olson and Hasegawa 1979, 1996; Olson 1980). Structure of the pelvis and legs is similar to that of other penguins, suggesting a marked upright stance, although the tarsometatarsus is longer than in later penguins. The femur is long and straight, as in other flightless wing-propelled divers, unlike the short and often-bent femur of foot-propelled divers such as cormorants, loons, and Hesperornis (see Miller and Howard 1949; Howard 1966, 1970, 1976; Olson and Hasegawa 1979, 1996; Olson 1980; Arney and Wise 2003). A long narrow bill occurs in other stem-penguins such as “Palaeeudyptes” and Platydyptes. The larger species (one specimen) is older at 61 Ma (see Supplementary Material); the smaller species (three specimens) is younger, approximately 58 Ma.
Evaluation of the Phylogenetic Position of Waimanu
In such cases, where a disparate new taxon is reported close to the K/T boundary, it should be considered whether the material represents a relict of a much older clade. In this case, Waimanu clearly shows synapomorphies for Ornithurae and Carinatae (characters discussed by Cracraft 1988) and, as indicated below, is deeply nested within the crown birds, Neornithes (see supplementary fig. 3).
We evaluated the phylogenetic position of Waimanu within Neornithes by cladistic analysis with the published data set of Mayr and Clarke (2003) with additional taxa but no new characters. We include three new taxa (Waimanu and the fossil penguins Platydyptes and “Palaeeudyptes”), giving 46 ingroup and 3 outgroup taxa, and 148 characters. Characters of Platydyptes and “Palaeeudyptes” were scored from specimens from the OU collections (Fordyce and Jones, 1990, T. Ando, 2006, PhD in progress). The data set was processed with PAUP* 4.0b10 (Swofford 2001) with the same settings as the primary analysis of the original analyses (Mayr and Clarke 2003) including bootstrap analysis.
Results show that Waimanu belongs to Sphenisciformes, namely to crown + stem-penguins sensu Clarke et al. (2003). In contrast to the molecular results reported here, the clade of loons and grebes (Gaviiformes + Podicipediformes) appears as the sister taxon to Waimanu + other penguins. Of note, Waimanu is not close to the Pelecaniformes or Charadriiformes which, according to other studies (Howard 1976, Olson and Hasegawa 1996), include wing-propelled diving clades: the extinct Plotopteridae (Pelecaniformes) and the extinct flightless auks in the Alcidae (Charadriiformes), see Supplementary Material.
The position of Waimanu is well supported. Unambiguously optimized synapomorphies place it in the Neornithes, the Neognathae, the Neoaves, the clade of Gaviidae + Podicipedidae + Sphenisciformes + Procellariidae, and the Sphenisciformes. Nodes for Neoaves and for the clade of Gaviidae + Podicipediformes + Sphenisciformes + Procellariiformes were not supported by the bootstrap analysis (50%), confirming the results of Mayr and Clarke (2003), but bootstrap analysis (50%) did support other clades including Sphenisciformes (see Supplementary Material Figure 4). Nodes and characters are as follows, with numbers in parentheses from the original data set of Mayr and Clarke (2003) (note that optimization has in some cases allocated state 0 as derived, and state 1 as primitive).
Neornithes: supratendinous bridge on distal end of tibiotarsus is completely ossified (100:1); distal interosseus canal is present in tarsometatarsus (107:0).
Neognathae: frontoparietal suture is closed (32:1); fossa of brachialis muscle of humerus is indistinct (79:1); scapulotricipital groove of humerus is well developed (81:1); ilioischiadic foramen of pelvis is closed posteriorly (94:1).
Neoaves: preacetabular tubercle of pelvis is vestigial (93:1).
Clade of Gaviidae + Podicipediformes + Sphenisciformes + Procellariiformes: nasal gland fossa is present on dorsal surface of frontal (25:1); pneumatic foramen is absent from bottom of pneumotricipital fossa of humerus (77:0); ulna does not distinctly exceed humerus in length (82:0); cnemial crest markedly protrudes proximally (99:1).
Sphenisciformes: some thoracic vertebrae are not heterocoelous (57:0); synsacrum has 11–12 ankylozed vertebrae (91:1); hypotarsal crests and grooves of tarsometatarsus are not well developed (103:0).
Mitochondrial Genomes and Phylogenetic Analysis
DNA extraction, long-range PCR, and subsequent rounds of short-range PCR (including cloning where necessary) and DNA sequencing were by standard methods and are reported in Supplementary Material. Accession numbers and sequence lengths for albatross, petrel, and loon are AY158677 (17,026 bp), AY158678 (16,414 bp, control region incomplete), and AY293618 (17,573 bp), respectively. Names and accession numbers of the other taxa analyzed are in Supplementary Material. The data set has 25 birds plus an outgroup of six reptiles, and consists of the 12 protein-coding genes from the heavy DNA strand together with 22 combined RNA genes. Alignments are at http://awcmee.massey.ac.nz/downloads.htm. Analysis is by standard programs including ModelTest (Posada and Crandall 1998), PAUP* (Swofford 2001), MrBayes (Huelsenbeck and Ronquist 2001), and Multidivtime (Thorne and Kishino 2002).
Phylogenetic analyses (see Supplementary Material) used nucleotide coding for first and second codon positions and for RNA stems and loops, and RY-coding for third codon positions. For both mammals and birds we find that increased taxon sampling (Lin et al. 2002) and RY-coding of third codon positions (Delsuc et al. 2003; Phillips and Penny 2003; Harrison et al. 2004), increases concordance between mitochondrial and nuclear data sets. We ran 1000 unconstrained ML bootstrap replicates with PAUP* on the Helix computing cluster (www.helix.massey.ac.nz), plus a Bayesian analysis using chains of 107 replicates. Detailed results are given in Supplementary Material. The reptilian outgroup joins between paleognaths and neognaths, consistent with recent work (Harrison et al. 2004). Of the outgroup taxa, crocodilians are closest to birds. Dating estimates using Multidivtime (Thorne and Kishino 2002) under a variety of constraints are also provided in Supplementary Material and show the high information content of the data. Due to program limitations of Multidivtime, divergence time estimation could not use RY-coding for the third codon position; therefore third positions were either omitted (as in fig. 2) or coded as nucleotides (as in Supplementary Material). The two main calibration points occur in different sections of the avian tree. We used 62 Ma for the divergence of penguins and storks (which are closest to penguins in the present mtDNA analysis) to allow time for divergence from their common ancestor. 66 Ma was used for the magpie goose/Presbyornis/Vegavis divergence (Kurochkin et al. 2002; Harrison et al. 2004; Clarke et al. 2005). Both calibration points are minimum divergence times on, for example, the penguin lineage. That is, they are estimates of the ‘lower bounds’ on the divergences, not estimates of the absolute dates. This is what is required to test the models of Penny and Phillips (2004). If the real divergences are older, this would only strengthen our conclusion about modern birds overlapping in time with earlier groups.
![Rooted tree for 25 birds showing posterior divergence time estimates and their standard deviations (95% confidence intervals were also calculated but are not shown). The tree is drawn to scale (time in million years [Myr]) and the Cretaceous/Tertiary (K/T) boundary at 65 Ma is marked. Dating estimates were carried out using the tree in Figure S2 (see Supplementary Material) and the program packages PAML and Multidivtime. The data set consisted of the 12 protein-coding genes from the mitochondrial heavy strand plus the two ribosomal RNAs and 20 transfer RNAs (tRNA-Phe and tRNA-Glu were excluded due to missing data), coded as nucleotide (nt) data. Due to limitations of Multidivtime, third codon positions (cdp) could not be analyzed as transversion (RY) data and were therefore omitted along with gaps, ambiguous sites around gaps and stop codons, giving a data set 8582 nt long. Two main calibration points were used (indicated in bold): 1. The divergence between magpie goose and (duck, goose) constrained at 66 Ma; 2. The divergence between penguin and stork constrained at 62 Ma. The F84 plus discrete gamma model was used in base_ml (PAML). Details of parameters used in are in Supplementary Material.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/mbe/23/6/10.1093_molbev_msj124/1/m_molbiolevolmsj124f02_lw.jpeg?Expires=1716340221&Signature=AoGKUCt55zvAdTL136DDP5PkG2M52DXtzLGpt8LYuYoAUg~qfJXPXjJ4K9A6obAicpoO8opI2gPmK6NY1pe~AVNT6B-5d-wUoLsDxV255MKRTI1E74TRkPsayZUMfsMUs2WP7aBQbq7x0iOZ2Vwqdrv1ofpqVyOX0I5QlPyC5-Eizeu9SdNpaN~bfE6oZyo0~sKKU4M-Q6bNFRo33Mk7Kce3xKJHKv622Ca7eHMjuCLJ7ptjLb1sNPFbZ717z2Gk8JPxPT4uFhM4MaW2vw6IBavNaF5oAJ0XBIBhMBOKkIDgHs91NOJGzosMmlRaahkoFlweC36OvjcDXI41deQFUw__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Rooted tree for 25 birds showing posterior divergence time estimates and their standard deviations (95% confidence intervals were also calculated but are not shown). The tree is drawn to scale (time in million years [Myr]) and the Cretaceous/Tertiary (K/T) boundary at 65 Ma is marked. Dating estimates were carried out using the tree in Figure S2 (see Supplementary Material) and the program packages PAML and Multidivtime. The data set consisted of the 12 protein-coding genes from the mitochondrial heavy strand plus the two ribosomal RNAs and 20 transfer RNAs (tRNA-Phe and tRNA-Glu were excluded due to missing data), coded as nucleotide (nt) data. Due to limitations of Multidivtime, third codon positions (cdp) could not be analyzed as transversion (RY) data and were therefore omitted along with gaps, ambiguous sites around gaps and stop codons, giving a data set 8582 nt long. Two main calibration points were used (indicated in bold): 1. The divergence between magpie goose and (duck, goose) constrained at 66 Ma; 2. The divergence between penguin and stork constrained at 62 Ma. The F84 plus discrete gamma model was used in base_ml (PAML). Details of parameters used in are in Supplementary Material.
Figure 2 shows the avian part of the tree, together with dating estimates. The combined chicken and duck group (Galloanseres) is basal within neognaths. The remaining neognath birds form Neoaves (Cracraft 2001), with the two clades represented here by the passerines and the ‘seabirds’/shorebirds/raptors. The division between shorebirds (turnstone and oystercatcher) and the informal group, which for present purposes we call seabirds (albatross, petrel, stork, loon, and penguin) is maintained (Cracraft 2001). The falconiforms (falcon and buzzard) are just basal to these. We do not, as reported earlier with short sequences (Cooper and Penny 1997), find the penguin joining with the procellariforms (petrel and albatross). The most unexpected aspect of the tree is the penguin/stork grouping, but we have omitted the stork, reanalyzed the data and still obtain the ((penguin, loon), (albatross, petrel)) grouping (see later). Thus the stork/penguin grouping does not affect our conclusions, but the relationships in that part of the tree are being addressed by sequencing of other potential relatives for both the stork and the penguin.
The dates of divergence shown in figure 2 are still conservative, but our results are consistent with the fossil record of bones and footprints. There are many fragmentary fossil bird remains in the Late Cretaceous (Hope 2002, Fountaine et al. 2005—and see http://palaeo.gly.bris.ac.uk/data/birds.html), together with a large body of fossil footprint data (Lockley 1998; Hwang et al. 2002; Lockley and Rainforth 2002). Because this material is fragmentary, certainly compared to the preservation of Waimanu, it is must be omitted from formal cladistic analyses and so the fossils are difficult to place taxonomically. Nevertheless, the birds existed and would have had ecological requirements and effects! A few new specimens (Kurochkin et al. 2002; Clarke et al. 2005) are important exceptions in that they are well preserved. Because our results come from a combination of excellent fossil material and long DNA sequences, they enhance the value of both these earlier fragmentary fossil bones and footprints.
There is wide debate about the identity and significance of Late Cretaceous Neornithes (modern birds). One recent review gave 44 Late Cretaceous records (Hope 2002, table 2); most are Maastrichtian, but six are Campanian and one (at 84–89 Ma) is Coniacian (giving a lower bound on the divergence of neognaths and paleognaths). Many were identified as members of stem lineages of current neornithine groups and, further, Robertson et al. (2004) gives a long list of Neornithes likely to have been present in the Late Cretaceous. Fountaine et al. (2005) recognized 22–23 species of Neornithes published by 2003 (database of Fountaine et al. 2005 at http://palaeo.gly.bris.ac.uk/data/birds.html; derived from Chiappe and Dyke (2002) and Hope's 2002 data). Fountaine et al. (2005) reiterated that the material is largely fragmentary, and indicated that while most specimens are of uncertain taxonomic affinities their ecological habitat may be more certain. Amongst the few Cretaceous neornithines known from reasonable material, Vegavis is based on one well-preserved associated skeleton (Clarke et al. 2005), and Teviornis is known from associated wing bones (Kurochkin et al. 2002). A third supposed neornithine, the Antarctic loon-like Polarornis, is of debatable age (Chiappe and Dyke 2002). In their overview, Fountaine et al. (2005) argued that neornithines really were rare in the Cretaceous, whereas Hope (2002) noted that 20–25 records from the Lance formation are neornithines. For now, our approach is to combine excellent fossil material from the early Paleocene close to the K/T boundary, with long DNA sequences to emphasize that the neornithines radiated in the Late Cretaceous. Our results show the advantage of combining fossil and sequence data.
The Cretaceous radiation of neornithines raises the question of competition, before the K/T boundary, between modern birds and archaic birds and pterosaurs. Competitive interaction could lead to ecological displacement, but the groups under study must have at some point overlapped in space and time and had ecological overlap. Major differences in body size would seem to rule out ecological overlap between any two clades, although disparate size might reflect earlier competitive divergence. If there is a long-term drop in diversity, the time of extinction of the last member in the clade is not as important (in ecological terms) as the preceding pattern of decline (Penny and Phillips 2004). Finally, it must be asked whether low-diversity clades might still be ecologically significant on a regional to global scale.
To consider other Cretaceous birds, the enantiornithines formed the most diverse clade of Mesozoic birds, and are structurally similar to later Neornithes (e.g. Zhou 2004). Enantiornithines apparently did not survive the KT boundary. The enantiornithine stratigraphic record, according to data from Fountaine et al. (2005, http://palaeo.gly.bris.ac.uk/data/birds.html), shows two diversity peaks in the Early and Late Cretaceous (Barremian-Albian and Campanian-Maastrichtian). Although there is an expanding Cretaceous record of footprints (e.g. Kim et al. 2003; Lockley 1998), such traces generally are only identifiable to a higher-level group such as bird or pterosaur, or “ecological” group such as “web-footed bird”. The results in figure 2 imply that shorebirds were present in the Campanian and Maastrichtian, and so they are the best candidates for leaving the “web-footed” footprints.
Decline in Pterosaur Diversity
Fossil sites with thousands of footprints (Kim et al. 2003) show that large pterosaurs lived in the same habitat as web-footed birds, although the differences in size, structure, and form of locomotion point to different ecologies for pterosaurs and birds. Wang et al. (2005) suggest that earlier (Aptian) pterosaurs were common. Kim et al. (2003) offer the interpretation that pterosaurs might have fed on small birds—equally plausible to us, however, is that the raptors (Falconiformes; which, as shown in fig. 2 are an early group) could have preyed on young pterosaurs. There are fundamental trends in the long-term record of pterosaurs (fig. 3).
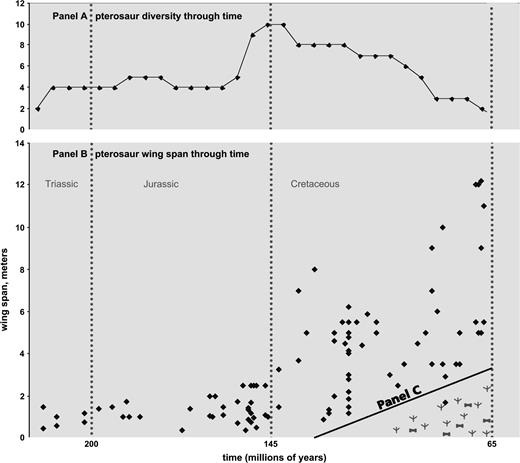
Pterosaurs through time. (A) Pterosaur taxonomic diversity from the mid-Triassic until the end of the Cretaceous (calculated at 5 Ma intervals from Fig 21 in Unwin, 2003). (B) Adult wingspans for pterosaurs. Estimates, wherever possible, are for adults and toward the larger size; sources are in Supplementary Material. The results indicate a major increase in size throughout the Cretaceous but with an increasing loss of small and medium-sized pterosaurs in the Late Cretaceous—at the same time modern birds are diversifying. (C) Occurrence of fossil footprints and stem-group modern birds in the last quarter of the Cretaceous. The panel is an indication that both fossil footprints of unidentified modern birds, as well as fossils of stem lineages, do occur in the last half of the Late Cretaceous.
The first aspect of pterosaur diversity is ecological/life history (fig. 3B) as revealed by trends in wingspan (see Wellnhofer 1991; Buffetaut and Mazin 2003; Chang et al. 2004; and further details in Supplementary Material). From the Late Triassic (Norian, ∼225 Ma) until the end of the Jurassic (∼145 Ma), the wingspan of pterosaurs was basically less than 2 m (meters), though with an increase toward the end of the Jurassic. From the early Cretaceous onward there was a major increase in maximum wingspan reaching to over 11 m by the end of the Cretaceous. Perhaps critical in relation to bird evolution was the loss of small pterosaurs (<2 m wingspan) from the mid-Cretaceous onward, matching the inferred radiation of Neornithes. (The only exception is a fragmentary record of Ornithocheirus bunzeli from 1881 which was <2 m and that needs to be confirmed; see Wellnhofer 1991.) The matter of size is complicated because many pterosaurs had distinct age classes, so that some previously described small ‘species’ were younger age classes of other species (see Bennett 1996).
The second aspect of pterosaur diversity is taxonomic. Panel A shows the highest diversity of pterosaurs around the end of the Jurassic/early Cretaceous; the later drop in pterosaur diversity matches the inferred radiation of Neornithes. These results are taken from figure 21 in Unwin (2003, shown also in Supplementary Material), except that we are not yet able to confirm records of nyctosaurids in the Maastrichtian, and thus terminate that lineage earlier. On quantitative grounds, the azhdarchids are by far the most common form in the Maastrichtian. The conclusion from Panels A and B is that, because the pterosaurs are reducing in diversity over the Late Cretaceous, the impact at the K/T boundary (even if it were the proximate cause) cannot be the ‘ultimate’ cause of pterosaur decline and eventual extinction.
The final small panel (C) is a qualitative reminder that both fossil remains of stem lineages of modern shore birds (Hope 2002) and fossil footprints (e.g. Lockley 1998; Hwang et al. 2002) exist during the later stages of the Cretaceous. The panel illustrates that from cause and effect considerations, modern shorebirds existed before the disappearance of pterosaurs. Similarly, we have reported (Fig. 3 in Penny and Phillips 2004) the decline in small dinosaurs (less than about 2 m long as adults) before the end of the Cretaceous. This decline in both small dinosaurs and pterosaurs is significant because it is different to the current situation in which it is the larger mammals, not the smaller, that are endangered (Cardillo et al. 2005). It is the larger species (not the smaller) that are at most risk of extinction. One inference from this is that pterosaurs were particularly vulnerable to extinction with only large species surviving.
Discussion
The four Waimanu fossils are some of the oldest well-preserved Neornithine birds. They pre-date by 6–8 Myr the widely cited Early Eocene species from the London Clay (53–55 Ma) (Feduccia 1996, 2003; Chiappe and Dyke 2002; Dyke and Gulas 2002), and are only a little younger than the best-associated partial skeleton of a neornithine, that of Vegavis (Clarke et al. 2005) from the Late Cretaceous. Waimanu indicates that penguins diverged from other Neornithes and acquired disparate features such as large body size, more upright stance, and wing-propelled diving habits by the early Paleocene—only 3–4 Myr after the K/T boundary event. We suggest that the great disparity between Sphenisciformes and their sister-taxa is consistent with an origin for penguins during the Late Cretaceous neornithine radiation. The time of radiation, which has been debated widely (Cooper and Penny 1997; Bleiweiss 1998; Cracraft 2001; Chiappe and Dyke 2002; Feduccia 2003), is predicted here as starting at 90–100 Ma. Our results do not yet fully resolve ingroup relationships for the seabird clade (see Cracraft 1988; Sibley and Ahlquist 1990; Cooper and Penny 1997; Livezey and Zusi 2001; Mayr and Clarke 2003).
Our results support the recent consensus that modern birds were not restricted to shorebird/seabird niches during the Late Cretaceous (Hope 2002; Dyke and van Tuinen 2004; but see Feduccia 2003). Both morphology (Cracraft 2001) and molecules place the root of the avian tree distant from the marine groups studied here; neither shorebirds nor seabirds are basal among modern birds. Because of the increased chance of finding fossils from an aquatic environment it is easy to under-estimate the relative age of non-aquatic birds (Hope 2002). The fossil record for aquatic birds appears reasonable (Fountaine et al. 2005) but, as judged from figure 2, most terrestrial lineages are not represented in the early fossil record. The best terrestrial bird record often comes from occasional settings of exceptional preservation (Norell and Clarke 2001; Zhou 2004).
A Late Cretaceous radiation of modern (neornithine) birds prompts questions about the evolution and extinction of earlier stem-birds (such as the enantiornithines, Hesperornis, and Ichthyornis) and pterosaurs. Were these groups displaced by modern (neornithine) birds, or did they replace the archaic groups after the latter disappeared? It is not clear whether the extinction of archaic birds was abrupt or involved long-term decline over some 30 Myr of the Late Cretaceous (Unwin 1988; Chiappe and Dyke 2002; see also Ward et al. 2005), when diverse crown-lineage birds were already present. Fossil footprints indicate that pterosaurs certainly coexisted with web-footed birds around 80–96 Ma (Hwang et al. 2002; Kim et al. 2003) (Campanian-Cenomanian), implying that some specialized birds occupied fresh-water settings in the Late Cretaceous. Some archaic forms clearly survived quite late—the flightless marine bird Hesperornis until early Maastrichtian (Hills et al. 1999), and Ichthyornis-like birds until the latest Cretaceous (Dyke et al. 2002; but debated by Clarke 2004). Following Penny and Phillips (2004) our focus is on when a group started to decline, not when the last member of a clade went extinct (which is intrinsically uncertain because of sampling issues from the Signor-Lipps effect [Wagner 2000]). Because archaic and modern birds overlapped for many million years, we focus on the ecological implications of this. When, for example, did modern birds start displacing/replacing earlier groups such as ichthyornithids; when did the earlier groups start losing niche-space to modern birds (Penny and Phillips 2004)?
Models for potential interactions between modern birds, archaic birds, and pterosaurs require life history information. For example, early pterosaurs show both slow bone growth and year classes (Bennett 1996), thus they may have been K-selected. Early birds, such as enantiornithines, show comparable patterns, with bone having distinct lines of arrested growth (Chinsamy et al. 1998). In contrast, Hesperornis and Ichthyornis, like neornithines, had rapid and sustained bone growth (Chinsamy et al. 1998), suggesting that later bird clades were physiologically advanced over earlier lineages. Given the recent focus (Holdaway and Jacomb 2000; Johnson 2002; Cardillo et al. 2005) on the susceptibility to extinction of large, K-selected animals (with slow growth and reproductive rates), life history patterns are important. Traditionally, there has been an apparent conflict between paleontological and molecular estimates of divergences for both mammals and birds. In contrast, we see important interactions between molecular and paleontological information; collaborative work is important. We need to integrate studies on paleontology (including footprints), DNA sequences (nuclear and mitochondrial), ecology, and physiology to develop testable models of past extinctions and radiations. Macroevolutionary models can then lead to testable models (Penny and Phillips 2004). One of the simplest predictions is an increase in the size of fossil pterosaur footprints during the Late Cretaceous, at the same time fossil footprints occur of shore birds. Similarly, we might expect the food sources of pterosaurs (as indicated by aspects of the fossils themselves, see Wellnhofer 1991) to decline in the Late Cretaceous. More predictions and tests are required.
K.E.S., C.M.J., and T.A. contributed equally to this work.
William Martin, Associate Editor
We thank Dick Gill (Department of Conservation Waikanae office) for the petrel sample, Alan Tennyson and Sandy Bartle of Te Papa (Museum of New Zealand) for the albatross, Einar Arnason and Thorvaldur Björnsson [Reykjavik, Iceland] for the loon, Trish McLenachan, Alicia Gore, and Gillian Gibb for some laboratory work, Matt Phillips and Fred Delsuc for assistance with analysis, David Unwin and Peter Wellnhofer for advice on pterosaur evolution, Gerald Mayr for providing a cladistics matrix for birds, Graeme Wilson for dinoflagellate dating, Norton Hiller for allowing study of the Canterbury Museum material, Te Papa for the use of facilities, and the Massey interloan service for assistance beyond the call of duty. The NZ Marsden Fund, the Otago Research Committee and the Swedish Science Council provided financial support. K.E.S. and G.L.H. carried out the DNA work; C.M.J., T.A., and R.E.F. the fossil part of the study (including naming and describing of Waimanu and both its species), T.A. the cladistic analysis of fossils, K.E.S. the phylogenetic and dating analysis, D.P. the pterosaur analysis, D.P. and U.A. designed the project, and D.P. and K.E.S. developed the first draft. We thank the landowners (Forrester and Goorde families) for access to localities. Finally, we thank Brad Field for finding the first specimen (OU 12651), and we particularly acknowledge the sterling efforts of Al Mannering in finding and preparing the Canterbury Museum specimens.
Funding to pay the Open Access publication charges for this article was provided by the Allan Wilson Center for Molecular Ecology and Evolution.
References
Alvarez, L. W., W. Alvarez, F. Asaro, and H. V. Michel.
Arney, J. E., and S. W. Wise Jr.
Baker, A. J., S. L. Pereira, O. P. Haddrath, and K.-A. Edge.
Bennett, S. C.
Bleiweiss, R.
Bromham, L. D., and M. D. Hendy.
Buffetaut, E., and J.-M. Mazin.
Cardillo, M., G. M. Mace, K. E. Jones, J. Bielby, O. R. P. Bininda-Emonds, W. Sechrest, C. D. L. Orme, and A. Purvis.
Chang, M-M., ed.
Chiappe, L. M.
Chiappe, L. M., and G. J. Dyke.
Chinsamy, A., L. D. Martin, and P. Dodson.
Clarke, J. A.
Clarke, J. A., and M. A. Norell.
Clarke, J. A., E. B. Olivero, and P. Puerta.
Clarke, J. A., C. P. Tambussi, J. I. Noriega, G. M. Erickson, and R. A. Ketchman.
Conway Morris, S.
Cooper, A., and D. Penny.
Cracraft, J.
Cracraft, J.
Cracraft, J., and J. A. Clarke.
Delsuc, F., M. J. Phillips, and D. Penny.
Dyke, G. J., R. W. Dortangs, J. W. M. Jagt, E. W. A. Mulder, A. S. Schulp, and L. M. Chiappe.
Dyke, G. J., and B. E. Gulas
Dyke, G. J., and M. van Tuinen.
Elzanowski, A., G. S. Paul, and T. A. Stidham.
Field, B. D., and G. H. Browne.
Foote, M., J. P. C. Hunter, M. Janis, and J. J. Sepkowski.
Fordyce, R. E., and C. M. Jones
Fountaine, T. M. R., M. J. Benton, G. J. Dyke, and R. L. Nudds.
Fox, R. C.
Galton, P. M., and L. D Martin.
Härlid, A., A. Janke, and U. Arnason.
Harrison, G. L., P. A. McLenachan, M. J. Phillips, K. E. Slack, A. Cooper, and D. Penny.
Hedges, S. B., P. H. Parker, C. G. Sibley, and S. Kumar.
Hills, L. V., E. L. Nicholls, L. M. Nunez-Betelu, and D. J. McIntyre.
Holdaway, R. N., and C. Jacomb.
Hollis, C. J., and C. P. Strong.
Hope, S.
Howard, H.
Howard, H.
Howard, H.
Huelsenbeck, J., and F. Ronquist.
Hwang, K.-G., H. Huh, M. G. Lockley, D. M. Unwin, and J. L. Wright.
Johnson, C. N.
Jones, C. M., and A. Mannering.
Kim, C. B., M. Huh, C. S. Cheong, M. G. Lockley, and H. W. Chang.
Kurochkin, E. N., G. J. Dyke, and A. A. Karhu.
Lin, Y.-H., P. A. McLenachan, A. R. Gore, M. J. Phillips, R. Ota, M. D. Hendy, and D. Penny.
Livezey, B. C., and R. L. Zusi.
Lockley, M. G., and E. C. Rainforth.
Martin, L. D., and J. D. Stewart.
Mayr, G., and J. Clarke.
Miller, L., and H. Howard.
Myrcha, A., P. Jadwiszczak, C. P. Tambussi, J. I. Noriega, A. Gazdzicki, A. Tatur, and R. A. del Valle.
Norell, M. A., and J. A. Clarke.
Olson, S. L.
Olson, S. L., and Y. Hasegawa.
Olson, S. L., and Y. Hasegawa.
Paton, T., O. Haddrath, and A. J. Baker.
Penny, D., and M. J. Phillips.
Peters, S. E.
Phillips, M. J., and D. Penny.
Posada, D., and K. A. Crandall.
Robertson, D. S., M. C. McKenna, O. B. Toon, S. Hope, and J. A. Lillegraven.
Sibley, C. G., and J. E. Ahlquist.
Simpson, G. G.
Slack, K. E., A. Janke, D. Penny, and U. Arnason.
Swofford, D. L.
Thorne, J. L., and H. Kishino.
Unwin, D. M.
Unwin, D. M.
Vajda, V., J. I. Raine, and C. J. Hollis.
Van Tuinen, M., and S. B. Hedges.
Wagner, P. J.
Wang, X., A. W. A. Kellner, Z. Zho, and D. S. Campos.
Ward, P. D., J. Botha, R. Buick, M. O. De Kock, D. H. Erwin, G. H. Garrison, J. L. Kirschvink, and R. Smith.
Wilf, F., K. R. Johnston, and B. T. Huber.
Author notes
*Allan Wilson Center for Molecular Ecology and Evolution, Institute of Molecular BioSciences, Massey University, Palmerston North, New Zealand; †Division of Evolutionary Molecular Systematics, Department of Cell and Organism Biology, University of Lund, Solvegatan 2, S-223 62 Lund, Sweden; ‡Institute of Geological and Nuclear Sciences, Lower Hutt, New Zealand; and §Department of Geology, University of Otago, Dunedin, New Zealand

{kind=link}
{kind=link}
{kind=link}