



Abstract
Recent neuroimaging evidence indicates neural mechanisms that support transient improvements in creative performance (augmented state creativity) in response to cognitive interventions (creativity cueing). Separately, neural interventions via tDCS show encouraging potential for modulating neuronal function during creative performance. If cognitive and neural interventions are separately effective, can they be combined? Does state creativity augmentation represent “real” creativity, or do interventions simply yield divergence by diminishing meaningfulness/appropriateness? Can augmenting state creativity bolster creative reasoning that supports innovation, particularly analogical reasoning? To address these questions, we combined tDCS with creativity cueing. Testing a regionally specific hypothesis from neuroimaging, high-definition tDCS-targeted frontopolar cortex activity recently shown to predict state creativity augmentation. In a novel analogy finding task, participants under tDCS formulated substantially more creative analogical connections in a large matrix search space (creativity indexed via latent semantic analysis). Critically, increased analogical creativity was not due to diminished accuracy in discerning valid analogies, indicating “real” creativity rather than inappropriate divergence. A simpler relational creativity paradigm (modified verb generation) revealed a tDCS-by-cue interaction; tDCS further enhanced creativity cue-related increases in semantic distance. Findings point to the potential of noninvasive neuromodulation to enhance creative relational cognition, including augmentation of the deliberate effort to formulate connections between distant concepts.
Introduction
An accomplished creative innovator, Steve Jobs reflected that innovators sometimes “feel a little guilty” about taking credit for their creative achievements (Wolf 1996, February). “That's because they didn't really do it, they just saw something. Creativity is just connecting things,” and creative ideas occur when people “are able to connect …and synthesize new things.” Indeed, creativity research indicates that new creative ideas often take the form connections between concepts that had previously seemed far apart (Holyoak and Thagard 1995). Relational cognition that reveals distant connections, especially in the form of analogical reasoning, is frequently a mechanism for generating creative leaps in the sciences, arts, education, and industry (Bassok and Holyoak 1989; Holyoak and Thagard 1995; Costello and Keane 2000; Dunbar and Blanchette 2001; Dahl and Moreau 2002; Schunn et al. 2006; Chan et al. 2012; Pollack 2014). There has long been interest in improving relational cognition (Gick and Holyoak 1980, 1983), and current widespread recognition of the value of creative insights in the innovation economy (IBM 2010; NSF 2011, 2013; Harrison et al. 2012; Pollack 2014) makes efforts to boost creative relational cognition particularly timely.
Creativity is a broad and multiplex construct. As we, among many, have argued (Finke et al. 1992; Green 2016), creativity research is most instructive when it seeks to parse out a particular creative process for close investigation rather than seeking to engage creativity as a whole. The influential “creative cognition” approach (Finke et al. 1992; Ward et al. 1995, 1999) posits that the most promising target processes for creativity research are the same processes that are cornerstones of complex cognition more broadly. In this respect, relational cognition—and especially analogical reasoning—is an appealing target for investigation. Understanding the relatedness of experiences, ideas, objects, words, etc. is fundamental to successful cognition (Hofstadter 2001; Halford et al. 2010), and the emergence of more abstract relational thinking in place of more superficial featural similarity is a hallmark of human cognitive development (Gentner 1988). Rich theory and empirical work (Sternberg 1977; Gick and Holyoak 1980; Gentner 1983; Morrison et al. 2004; Bunge et al. 2005; Green, Fugelsang, and Dunbar 2006; Green, Fugelsang, Kraemer, et al. 2006; Green et al. 2008) has further parsed relational cognition into subprocesses at both the cognitive and neural levels, enabling a sharper focus for targeting constrained elements of creative cognition. A specific component process of relational cognition that has been a fruitful target for creativity research is the process of relational integration (Green et al. 2010; Green, Kraemer, et al. 2012), which refers to the formation of a unified mental representation of a relation by integrating separate semantic elements (e.g., forming a unified representation of second-order relation by integrating multiple first-order relations in analogical reasoning). Relational integration is cognitively and neurally distinguishable from (though largely overlapping with) other semantic processing components of relational cognition such as retrieval and alignment (Green, Fugelsang, and Dunbar 2006; Green, Fugelsang, Kraemer, et al. 2006; Green et al. 2008). Understanding how relational integration works and whether it can be bolstered has obvious implications for creative insights that rely on distant connections. However, consistent with the creative cognition approach, relational integration is not only relevant to creativity but also appears to support reasoning and decision-making functions more globally (Christoff et al. 2001; Ramnani and Owen 2004; Koechlin and Hyafil 2007; Shamosh et al. 2008; Boorman et al. 2009; Halford et al. 2010).
Neuroimaging research has provided convergent implications that an anterior prefrontal brain region, frontopolar cortex, supports relational integration in service of analogical reasoning (Christoff et al. 2001; Bunge et al. 2005; Green, Fugelsang, Kraemer et al. 2006; Green et al. 2010; Green, Kraemer, et al. 2012), operating in concert with temporal, parietal, and other prefrontal cortical regions (Christoff et al. 2001; Morrison et al. 2004; Bunge et al. 2005; Geake and Hansen 2005; Green, Fugelsang, Kraemer, et al. 2006; Krawczyk et al. 2008; Wendelken et al. 2008; Cho et al. 2009; Green et al. 2010; Volle et al. 2010; Hampshire et al. 2011; Bassok et al. 2012; Green, Kraemer, et al. 2012; Knowlton et al. 2012; Watson and Chatterjee 2012). Notably, relational integration of unified representations from disparate semantic elements in frontoplar cortex appears to cohere closely with the process of mapping unit representation of abstract, integrated concepts in frontopolar cortex within an elegant neurocomputational architecture for relational reasoning proposed by Knowlton et al. (2012). Indeed, several generally convergent models of prefrontal cognitive architecture indicate a rostro-caudal hierarchy in which frontopolar cortex integrates relatively less abstract pieces of information to form more highly abstract representations (Christoff and Gabrieli 2000; Ramnani and Owen 2004; Badre and D'Esposito 2007; Knowlton et al. 2012). Work in our laboratory has found that increasing frontopolar activity parametrically reflects increasing creativity in analogical reasoning, where creativity is measured quantitatively by “semantic distance” between the concepts that analogies connect (Green et al. 2010; Green, Kraemer, et al. 2012). This may be because integrating representations that are more disparate (i.e., more distant from each other in semantic space) places greater demand on frontopolar integration circuitry (Green, Kraemer, et al. 2012; Green et al. 2015); for review, see Green (2016).
Recent evidence indicates that consciously directed effort to think more creatively yields more semantically distant connections in analogy and simpler forms of relational cognition (Green, Cohen, et al. 2012; Green et al. 2015). This “thinking cap” effect (i.e., trying and succeeding at being more creative) is commonly experienced, and the recent findings in relational cognition accord with prior findings in other areas of creative performance (Gilchrist and Taft 1972; Harrington 1975; Runco and Okuda 1991; Seger et al. 2000; Niu and Sternberg 2001; O'Hara and Sternberg 2001; Chen et al. 2005; Howard-Jones et al. 2005). Nonetheless, the potential of the thinking cap effect remains greatly underexplored by creativity research, which has traditionally focused instead on creativity as a static trait (Guilford 1950, 1967; Mayer 1999). Intriguingly, we recently observed that conscious augmentation of performance on a creative relational cognition task was associated with changes in activity and connectivity of frontopolar cortex (Green et al. 2015). Specifically, the magnitude of increase in frontopolar activity predicted individual differences in the extent of increases in quantitative semantic distance between words used to form relations. This suggests the possibility that neural intervention to potentiate increases in frontopolar activity might facilitate augmentation of state creativity with respect to the formation of distant semantic connections; that is, potentiation of frontopolar neurons might enhance the capacity of an individual to deliberately become a more semantically distant thinker.
Separately from research on the thinking cap effect, direct neuromodulation via transcranial direct current stimulation (tDCS) has shown promise as another method to transiently augment creative performance (Cerruti and Schlaug 2009; Chi and Snyder 2011, 2012; Chrysikou et al. 2013; Simis et al. 2014; Brunye et al. 2015; Colombo et al. 2015; Goel et al. 2015; Mayseless and Shamay-Tsoory 2015). A timely question concerns the relationship between neuromodulation and deliberate/conscious augmentation of state creativity. Given the evidence that conscious, “thinking cap” state creativity augmentation is effective, including specific effects on relational cognition, can neuromodulation boost this effect further?
Another timely question about the potential of neuromodulation to augment creative cognition concerns analogical reasoning, which no neuromodulation study to our knowledge has yet addressed. The rich literature demonstrating the unique power of analogy for learning and innovating (Bassok and Holyoak 1989; Holyoak and Thagard 1995; Costello and Keane 2000; Dunbar and Blanchette 2001; Dahl and Moreau 2002; Schunn et al. 2006; Chan et al. 2012; Pollack 2014) suggests potentially far-reaching implications of augmenting analogy. At a construct level, analogical reasoning is an intriguing target for enhancement, because it bridges the divide between creativity and IQ (Holyoak 2012). Thus, if analogical reasoning can be improved via neuromodulation, this would support extending the boundaries of tDCS efficacy from creativity to a form of intelligence.
Finally, prior studies of the effects of cognitive and neural interventions on creativity have primarily focused on divergence (i.e., the novelty/unusualness of generated responses), whereas consensus criteria among creativity researchers require products to be both divergent and meaningful/appropriate to be considered truly creative; for review, see Runco and Jaeger (2012). While state creativity manipulations are promising, it is not yet clear whether these interventions represent “real” creativity or simply yield divergence by diminishing constraints on meaningfulness/appropriateness.
Here, we addressed these issues in 2 tasks of creative relational cognition, an open-ended analogical reasoning task (analogy finding task; Weinberger et al. 2016), and a modified verb generation task (thin slice creativity task; Prabhakaran et al. 2014). High-definition tDCS was used to potentiate neuronal activity of the specific frontopolar region in which increasing activity has been parametrically associated with increasing semantic distance in analogical reasoning (Green et al. 2010; Green, Kraemer, et al. 2012), and which has been shown to predict individual differences in state-related augmentation of creative performance in verb generation (Green et al. 2015). We tested the hypothesis that tDCS would interact with conscious augmentation of state creativity to produce even greater increases in semantic distance in the thin slice creativity verb generation task, using an explicit creativity-cueing paradigm with known efficacy and good construct validity in relation to conventional measures of creativity (Prabhakaran et al. 2014). Whereas anodal stimulation produces reliable facilitation of performance on cognitive tasks, cathodal stimulation produces more variable direction and magnitude of effects from study to study (Jacobson et al. 2012). We therefore utilized anodal tDCS only, which allowed a clear directional hypothesis of enhanced creativity with active tDCS compared with sham stimulation. Based on our neuroimaging findings (Green et al. 2010; Green, Kraemer, et al. 2012), we predicted that targeted frontopolar neuromodulation would bolster creative analogical reasoning and sought to test whether any observed increases in creativity reflected “real” creativity, rather than simply divergence at the expense of appropriateness. We employed a novel analogy finding task paradigm that was devised to be sufficiently open-ended to detect individual differences in creative analogical reasoning, yet sufficiently constrained to enable sensitivity to inappropriate responding (Weinberger et al. 2016).
Methods
Participants
Thirty-one right-handed, native English speakers (16 female, 15 male; average age = 21.69 ± SD = 3.09) were recruited via flyer advertisements on the Georgetown University campus and online via the Georgetown Research Volunteer Program and Research Match. Participants were randomly assigned to receive tDCS (N = 15) or sham (N = 16). tDCS and sham groups did not differ by age (tDCS: 22.50 ± 3.87 years; Sham: 20.98 ± 2.08 years; P = 0.183), sex (tDCS: 6 female; Sham: 10 female; P = 0.210), years of education (tDCS: 16.29 ± 2.05 years; Sham: 15.13 ± 2.06 years; P = 0.135), parents' highest level of education (P = 0.155), household income (tDCS: $75 000 ± $31 132; Sham: $80 000 ± $33 267; P = 0.675), verbal IQ derived from subscales of the Multiple Aptitudes Battery-II (MAB-II; Jackson (1998); tDCS: 119.07 ± 13.53; Sham: 119.64 ± 7.95; P = 0.893), mental rotations test accuracy (MRT; Vandenberg and Kuse 1978; Peters et al. 1995; tDCS: 48.81 ± 23.48%; Sham: 39.29 ± 18.77%; P = 0.246), or self-reported SAT/ACT scores (Cole and Gonyea 2010; tDCS: 2080.00 ± 240.28; Sham: 2103.57 ± 173.37; P = 0.768). All participants gave written, informed consent in accordance with the Georgetown University Institutional Review Board, which approved all procedures. Data for 5 participants (1 tDCS; 4 sham) were lost from the thin slice creativity verb generation task due to a data collection software error. One tDCS participant's data were lost from the analogical reasoning due to data collection software error. Two participants (1 tDCS; 1 sham) were excluded from the analogy finding task analysis because of extremely brief amounts of time taken to complete a task matrix (<20 s), which was deemed to indicate inappropriate attention devoted to task performance. Remaining group sizes nonetheless afforded 80% power to detect an interaction of creativity cue by neuromodulation in a planned mixed ANOVA model at a modest effect size of η2 = 0.078 with α = 0.050. Group sizes were also comparable with or greater than group sizes used in recent studies that have detected effects of tDCS on creativity (Cerruti and Schlaug 2009; Chrysikou et al. 2013; Brunye et al. 2015; Colombo et al. 2015; Mayseless and Shamay-Tsoory 2015). The measures of verbal IQ, MRT, and self-reported SAT were obtained via an online follow-up survey as we described previously (Ihne et al. 2016; Weinberger et al. 2016; see Supplementary Methods).
Neuromodulation
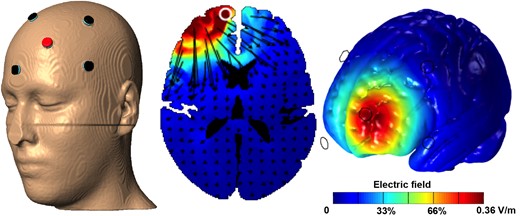
{kind=link}
HD-tDCS electrode montage and electrical field model. Electrical field modeling was performed in HD-Explore 3.1a (Soterix Medical, Inc.) using the MNI-152 template. The left panel shows the electrode locations rendered on the template head (red = anode, black = cathode). The middle panel shows an axial slice at the level of the maximal field. Black arrows depict the directional current flow vectors. The right panel depicts the total electrical field magnitude rendered on the template brain surface. Renderings were provided courtesy of Chris Thomas and Dr Abhishek Datta of Soterix Medical, Inc.
Tasks Procedures
Participants received anodal tDCS or sham for the first 20 min of the testing session. During the first 5 min, participants filled out a brief demographics survey and then rested. Participants then completed the analogy finding task, which was up to 5 min. Because the analogy finding task was self-paced, participants finished at different times. To equate as much as possible the cognitive demands on participants during this window of the tDCS session, a filler task was used in which participants were asked to speak aloud word pairs that were analogically related to word pairs appearing on the screen. This filler task lasted from the end of the analogy finding task until a point 14 min and 40 s after the onset of tDCS, such that all participants participated in this task for at least 4 min and 40 s. Participants then completed the thin slice creativity verb generation task for exactly 16 min in total (5 mins and 20 s during tDCS and 10 min and 40 s after). tDCS stimulation for 20 min, the duration used in the present study, has been shown to induce cortical excitability shifts that are stable at least an hour after cessation of active stimulation (Nitsche and Paulus 2000; Nitsche et al. 2003). The creativity tasks were presented, using a Dell Optiplex 390 with a 21-inch LCD monitor with a 120 Hz refresh rate. The analogy finding task was presented from www.qualtrics.com, whereas the thin slice creativity verb generation task was presented using Eprime 2.0. Spoken responses for the thin slice creativity verb generation task were recorded using a digital video camera and transcribed offline.
Analogy Finding Task
Analogical reasoning was tested using a novel analogy finding task paradigm (Weinberger et al. 2016). The analogy finding task consisted of 2 matrices of word-pairs (Appendix A). Each matrix contained 5 word-pairs arranged in a column on the left side of the screen (stem pairs) and 20 word-pairs arranged in a row across the top of the screen (completion pairs). Participants were instructed as follows: “Your task is to make analogies by combining word-pairs on the left side of the grid with word-pairs along the top of the grid. Each word-pair should be read as ‘[Top Word] is to [Bottom Word]’ For example, ‘Helmet is to Head.’ Check the boxes to indicate when a word-pair from the top combines with a word-pair on the left to make a valid analogy.” “Please think creatively as you search for valid analogies. Some analogies may not be obvious right away, so be sure to look for abstract connections. However, only valid analogies should be listed, so don’t list analogies unless you can describe how the two word pairs are analogous.” Each stem pair could be combined with 3 or 4 completion pairs to form valid analogies, such that there were a total of 17 potential valid analogies that could be found within each matrix (i.e., valid analogies formed by combining one of the stem pairs with one of the completion pairs), and 83 potential word-pair combinations that yielded invalid analogies in each matrix. There was no restriction against combining the same completion pair with >1 stem pair and indeed each matrix included 1 completion pair that could be validly combined with 2 stem pairs. This was done to ensure that participants did not eliminate completion pairs from consideration, keeping the search space equivalently large throughout the task. For each stem pair of each matrix, the possible valid analogies represented a range of semantic distances. Varying the semantic distance of available analogical mappings in this way enabled us to discern individual differences in the creativity of the analogies participants formed. The requirement to form valid analogies while avoiding far more numerous invalid analogical connections enabled a sensitive measure of selectivity versus inappropriate responding. A pilot study (reported in Weinberger et al. (2016)) in 178 participants tested online via Amazon Mechanical Turk confirmed that analogies intended to be valid were identified far more frequently than analogies intended to be invalid, McNemar's within-subjects χ2 = 276.64, P < 0.001. Additionally, the validity of analogies was determined by domain experts, and analogy items were largely drawn from sets of stimuli used in previous full and pilot studies in our laboratory that obtained high rates of participant accuracy.
Thin Slice Creativity Verb Generation Task
We employed the thin slice creativity verb generation task, which we have described previously, and have validated in relation to a range of established creativity measures (Prabhakaran et al. 2014; Green et al. 2015). On each trial of the task, participants were presented with a noun on a computer screen (8 s duration; 250 ms ISI) and asked to say a verb that was related to the noun in any way. A brief tone marked the beginning of each trial. Of the 120 nouns presented, half (60) were shown in the Cued condition and the other half (60) in the Uncued condition. Conditions were indicated by the color in which the words were presented (green or purple), and we counterbalanced which color indicated which condition between subjects. For nouns presented in the Cued color, participants were instructed to “think creatively when coming up with your response.” Conditions were interleaved such that 2 trials of each condition were followed by 2 trials of the other condition. This design allowed us to investigate acute engagement of state creativity while minimizing set shifting and executive attention demands (e.g., from unpredictable or surprising shifts), which were not the intended targets of investigation. We have previously demonstrated that this design avoids confounds of set shifting (Green et al. 2015). We further counterbalanced 2 lists of noun prompts presented in the Cued versus Uncued conditions, crossed with the counterbalancing of the cue colors to yield a total of 4 counterbalancing cells.
The 2 counterbalanced lists of noun prompts were equated for word length (number of letters; t(118) = 1.22, P = 0.225) and average constraint, t(118) = −0.014, P = 0.989, where constraint refers to the extent to which the noun is associated with a single common verb associate (Barch et al. 2000). Constraint level was based on normative pilot data from testing in an independent sample (verb responses were obtained for each noun from 62–85 adults as part of a larger set of verb generation stimuli; no creativity cue manipulation was used in this pilot testing; see Prabhakaran et al. (2014)). For example, the noun prompt, “Pie,” was relatively high constraint because >75% of pilot participants generated the same verb response, “Eat,” whereas the noun prompt, “Dirt,” was relatively low constraint because no single verb response accounted for as much as 25% of responses. From a full list of 161 piloted nouns, the present study included the 60 highest constraint and 60 lowest constraint nouns. The sequence of high- versus low-constraint noun prompts was randomized. The first 40 noun prompts (presented during active tDCS) and last 80 noun prompts (presented after cessation of tDCS) did not differ in constraint (t(118) = 0.056, P = 0.955) or in word length (t(118) = 0.788, P = 0.432).
Calculation of Semantic Distance
Creativity of responding in both tasks was quantified using a measure of semantic distance derived via latent semantic analysis (LSA; Landauer and Dumais 1997; Landauer et al. 1998; Dumais 2004; http://lsa.colorado.edu). We used the topic space of “general reading up to first year college (300 factors)” and term-to-term comparison type. This measure of semantic distance corresponds to the cosine of the angle between vectors (corresponding, in our usage, to the terms of each analogy or the noun prompt and the verb response) within a given semantic space, which is derived through analyses of all of the contexts in which each word tends to be present or absent in that topic space (Landauer et al. 1998). Semantic distance determined via LSA is a reliable and construct-valid measure of creativity (Wolfe and Goldman 2003; Forster and Dunbar 2009; Green et al. 2010; Prabhakaran et al. 2014), and has previously been shown to be a quantifiable measure of creativity in analogical reasoning that correlates with subjective creativity ratings of analogies (Green et al. 2010). Outputs from LSA computations directly reflect semantic proximity rather than semantic distance, with values ranging from 0 to 1. Thus, for ease of interpretation, we subtracted the raw values from 1 to represent semantic distance and then multiplied by 10 so that values are whole numbers.
Results
Analogical Reasoning (Analogy Finding Task)
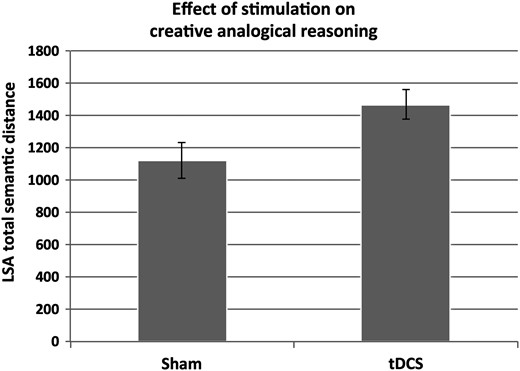
{kind=link}
Effect of tDCS versus sham on semantic distance in the analogy finding task. Anodal tDCS of frontopolar cortex was associated with the formation of more semantically distant analogical connections between word-pairs in the analogy finding task matrices. Error bars represent 1 SEM.
The total, rather than the average, semantic distance was used as the primary measure of creative analogical reasoning, because averages were deemed likely to provide misleading representations of the ability to formulate creative analogies. For example, it was possible to identify very few analogies (as few as one) and yet to have a high average semantic distance. Nonetheless, the analysis of average semantic distance also revealed an increase under tDCS (t(26) = 2.09, P = 0.046, d = 0.82). The total number of valid analogies identified, which is related but not informationally identical to total semantic distance, was also greater for tDCS (M= 20.69, SD = 3.72) relative to sham (M= 16.67, SD = 4.76, t(26) = 2.46, P = 0.021, d = 0.96). A nominally larger total number of responses (valid or not) under tDCS did not reach significance (t(26) = 2.00, P = 0.056, d = 0.78). Critically, the increase in semantic distance of analogy formation was not accompanied by an increase in the number of “false alarm” analogies formed under tDCS or in the number of false alarms as a proportion of the total number of analogies (both P > 0.385). Univariate ANCOVA comparing total semantic distance of valid analogies for tDCS versus Sham, with number of false alarms as a covariate, confirmed that the effect of tDCS on semantic distance was independent of false alarms (F1,25 = 4.67, P = 0.040, η2 = 0.16). The false alarms covariate was not significantly related to semantic distance in this model (F1,25 = 1.68, P = 0.206, η2 = 0.06). Indeed, the sensitivity index (d′) calculated as standardized false alarms subtracted from standardized valid analogies did not show a decrease under tDCS and actually showed a nominal though nonsignificant increase (t(26) = 1.15, P = 0.261, d = 0.45). tDCS (M = 231.15 s, SD = 21.89) was associated with nominally longer average time to complete the analogy finding task relative to sham (M = 194.86 s, SD = 68.86) though this difference did not reach significance, t(26) = 1.82, P = 0.081, d = 0.71). Longer time to complete the task is consistent with the evidence that participants produced a greater number of valid answers under tDCS. The analysis of false alarm analogies formed by participants indicated no effect of tDCS on total or average semantic distance (both P > 0.607), which suggests that tDCS may not have influenced the extent to which semantic distance/proximity influenced false choice selection.
Verb Generation (Thin Slice Creativity Task)
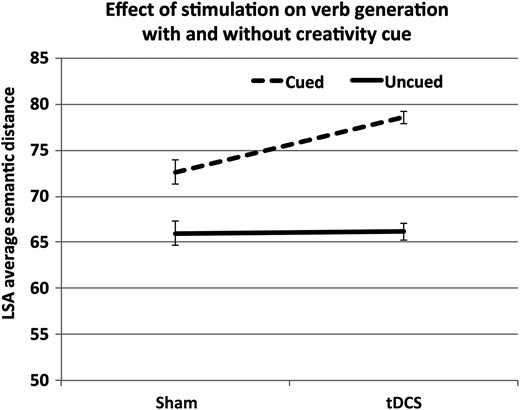
{kind=link}
Effect of tDCS versus sham (between-subjects) and Cued versus Uncued condition (within-subjects) on semantic distance of verb responses in the Thin Slices of Creativity task. Verb responses were uniformly more distant from noun prompts in the presence of the creativity cue. Anodal tDCS of frontopolar cortex augmented this effect, such that responding was most semantically distant when tDCS was combined with the creativity cue. Error bars represent 1 SEM.
To investigate effects of the constraint level of noun prompts, a within-subjects factor (Constraint: High, Low) was added to the ANOVA model. This model reconfirmed the main effects of Cue (F1,24 = 76.92, P < 0.001, η2 = 0.76); and Stimulation (F1,24 = 9.50, P = 0.005, η2 = 0.28); and the interaction of Cue by Stimulation (F1,24 = 96.22, P = 0.017, η2 = 0.22). Additionally, this model identified a main effect of Constraint (F1,24 = 96.22, P < 0.001, η2 = 0.80) indicating that, as expected, low-constraint noun prompts elicited more semantically distant verb responses; an interaction of Cue by Constraint (F1,24 = 10.79, P = 0.003, η2 = 0.31) indicating that constraint level had less effect on Cued noun prompts; an interaction of Stimulation by Constraint that approached but did not reach significance (F1,24 = 3.38, P = 0.078, η2 = 0.12), suggesting a greater effect of tDCS for high constraint noun prompts; and no 3-way interaction (F1,24 = 0.133, P = 0.718, η2 < 0.01). Confirmatory 2 × 2 models demonstrated significant interactions of Constraint by Cue (F1,25 = 11.05, P = 0.003, η2 = 0.31) and Constraint by Stimulation (F1,24 = 6.90, P = 0.015, η2 = 0.22).
The apparatus for recording spoken verb responses was separate from the stimulus presentation software, so reliable measures of response time were not readily obtained. Though response time was not an outcome measure of primary interest, an effort was made to reconstruct response times from videos of participants performing the thin slice creativity verb generation task. This reconstructed response time measure did not indicate main effects of Cue or Stimulation, or a Cue-by-Stimulation interaction (all P > 0.670).
Discussion
The present study addressed questions about the potential of cognitive and neural interventions to augment relational creativity (i.e., forming connections between distant concepts). An explicit creativity cueing paradigm with known efficacy (Green, Cohen, et al. 2012; Prabhakaran et al. 2014; Green et al. 2015; Weinberger et al. 2016) was used in 2 tasks of relational cognition, analogical reasoning, and modified verb generation. Creativity cueing was combined with high-definition tDCS targeted to potentiate activity in a frontopolar region that has been shown to reflect increasing semantic distance of relational integration in analogical reasoning (Green et al. 2010; Green, Kraemer, et al. 2012) and to predict the efficacy of conscious state creativity augmentation in thin slice creativity verb generation (Green et al. 2015). Data indicated that tDCS was effective in eliciting more creative (semantically distant) analogical reasoning without decreasing the meaningfulness/appropriateness of analogies, and that tDCS interacted with creativity cueing to facilitate more creative responding in verb generation.
To test state creativity effects on analogical reasoning, we employed a novel analogy finding task, which measures the ability to identify creative analogical connections in a large matrix of possible word-pair combinations. Analogy is a good model for creativity in reasoning because analogies can involve great divergence (highly creative analogies reveal hidden similarities between items or concepts that seem unrelated on the surface), but is also clearly constrained (to be valid, the pieces of an analogy must align with each other in meaningful/appropriate ways (Sternberg 1977; Gentner 1983; Hummel and Holyoak 1997)). These attributes of divergence and meaningfulness/appropriateness make analogy a form of reasoning that meets the modern consensus definition of creativity (for review, see Runco and Jaeger (2012)). Studies of creativity in analogical reasoning have generally focused on the evaluation of fully formed singleton analogies of the form “A is to B as C is to D,” or completion of nearly formed singleton analogies (Green, Cohen, et al. 2012; Vendetti et al. 2014). These paradigms greatly limit the search space in which reasoners seek and evaluate analogical connections. Valuable creative insights in real-world contexts cannot often be gained by simply assessing the validity of a fully formed analogy or adding a single term to complete a nearly formed analogy. Rather, creative analogical insights generally require the reasoner to seek out and find analogical connections that others may have missed among a broad array of possible concept combinations, where most of the possible combinations do not lead to valid analogies (Gick and Holyoak 1980, 1983; Holyoak and Thagard 1995; Costello and Keane 2000; Dunbar and Blanchette 2001; Dahl and Moreau 2002; Schunn et al. 2006; Chan et al. 2012). Thus, the much broader search space of the analogy finding task was designed to better inform understanding of the kind of reasoning that supports innovation. Because 83 of the 100 available connections between word-pairs in each analogy finding task matrix yielded invalid analogies, the task is highly sensitive to inappropriate responding. This enabled us to test whether putatively augmented state creativity in analogical reasoning might simply be due to participants decreasing the stringency with which they discern valid analogical connections (i.e., divergence at the expense of meaningfulness/appropriateness).
Comparison of tDCS with sham (both administered in the presence of the creativity cue) yielded 2 key results. First, tDCS elicited the formation of more creative (semantically distant) analogies. Second, this increase in the creativity of analogical reasoning was not due to an increase in invalid/false alarm analogies, and indeed false alarm analogies did not increase under tDCS. These findings provide evidence that tDCS can improve a valuable form of creative intelligence, analogical reasoning, which no tDCS study to our knowledge has explored.
Based on prior evidence, it is likely that neuromodulation of frontopolar cortex facilitated the relational integration function subserved by this region during analogical reasoning (Christoff et al. 2001; Bunge et al. 2005; Green, Fugelsang, Kraemer et al. 2006), especially facilitating more semantically distant relational integration that appears to place greater demand on frontopolar circuitry (Green et al. 2010; Green, Kraemer et al. 2012).
To explore the interaction between neural and cognitive intervention, we used a repeated, “on-off” creativity cue paradigm in the thin slice creativity verb generation task, which we have previously validated in relation to established measures of creativity (Prabhakaran et al. 2014). This paradigm provides a robust measure of the effect of cognitive state creativity intervention because an augmented creative state is dynamically engaged and disengaged multiple times in response to explicit cueing (60 times in the present study). As expected, the creativity cue led to increased semantic distance in verb generation in both the tDCS and sham groups. tDCS did not yield greater semantic distance than sham in the absence of the creativity cue. However, tDCS interacted with the creativity cue such that semantic distance was greater for tDCS than sham in the presence of the creativity cue. These findings appear to be consistent with recent evidence that the extent of activation increase in the frontopolar region we targeted predicts individual extent of increase in semantic distance during the thin slice creativity task (Green et al. 2015). If increasing frontopolar cortex is a neural mechanism that supports successful conscious augmentation of relational creativity (i.e., forming more distant relations between semantic representations), then potentiating frontopolar activity via anodal tDCS may facilitate this augmentation. Indeed, our data indicate that neuromodulation of frontopolar cortex may enhance the capacity to deliberately formulate more semantically distant relational connections.
The present experimental manipulation data converge with prior observational data from neuroimaging that have suggested a role for frontopolar cortex in relational integration (Christoff and Gabrieli 2000; Bunge et al. 2005; Green, Fugelsang, Kraemer, et al. 2006). Taken as a whole, this body of evidence concerning frontopolar cortex as an integrator of information to formulate abstract representations based on more caudally represented semantic elements appears well fitted to the general form of a rostro-caudal hierarchical model of prefrontal cognition that has emerged from the convergence of several important accounts (Christoff and Gabrieli 2000; Ramnani and Owen 2004; Badre and D'Esposito 2007; Knowlton et al. 2012). Ramnani and Owen (2004), identifying coherence between neurocognitive data and histological architecture, noted that extensive arborization relative to cell size in frontopolar neurons suggests wiring optimized for integration, particularly in service of abstract representations.
Knowlton et al. (2012) have proposed a neurocomputational account of relational reasoning (including analogy) in which a hierarchical progression of “role bindings” of semantic elements within a propositional information structure culminates in the abstract, integrated representation of similarities between propositions. The authors suggest that these highest level integrated representations (mapping units) are instantiated by neuronal activity in frontopolar cortex. Interpreted in this context, it may be that anodal tDCS in the present study facilitated frontopolar circuitry in accessing and/or integrating lower order role bindings. The account proposed by Knowlton and colleagues specifies that synchronization in the gamma and theta bands facilitates relational binding in the frontal lobes, and that cross frequency coupling between theta and gamma oscillations provides a mechanism for coordination between parietal storage areas and frontal relational processors, including the frontopolar cortex. Anodal tDCS has previously been shown to increase gamma power in hippocampal slice preparations (Reato et al. 2015), and both theta and gamma power when applied over the prefrontal cortex in humans (Miller et al. 2015). tDCS is thought to increase synchronization of brain networks connected to the directly stimulated area (Kunze et al. Forthcoming), and in some cases, anodal tDCS has been shown to increase theta coherence between the frontal and parietal lobes (Polania et al. 2011; Notturno et al. 2014). Thus, although we did not measure changes in theta or gamma oscillations here, our findings are potentially consistent with the account by Knowlton and colleagues, which predicts that enhancing frontoparietal theta coherence and prefrontal theta and gamma would augment the ability to draw more distant conceptual relationships. Additional research using alternating current stimulation to enhance oscillations at specific frequencies will be needed to confirm this hypothesis.
A worthy goal for future empirical and theoretical research is to characterize the interaction between extant hierarchical prefrontal neurocognitive architectures and emerging efforts to identify neural systems common to multiple forms of creativity. Promising brain-based accounts of creativity have emphasized connectivity between prefrontal executive cortical regions and the so-called default mode network of frontal and posterior midline regions (Jung et al. 2013; Beaty et al. 2016). These accounts ascribe an idea-generative function to default mode regions within the process of developing original thoughts, whereas prefrontal executive regions instantiate evaluative selection of ideas based on relevant constraints. It is unlikely that frontopolar cortex, a cortical region corresponding roughly to Brodmann area 10, can be grouped with the classical, task-negative default mode network (Raichle et al. 2001) given the extensive evidence that frontopolar activity shows task-positive responsivity across a range of cognitive domains including reasoning, economic decision-making, and branching/sub-goaling in addition to creativity (for relevant reviews, see Ramnani and Owen (2004); Gilbert et al. (2006); Koechlin and Hyafil (2007); Knowlton et al. (2012); Green (2016)), and that increasing task demand results in increased rather than decreased frontopolar activity (Koechlin and Hyafil 2007; Green 2016). The present data suggest that potentiating greater activity in frontopolar cortex facilitated both a primarily generative task (verb generation) and a primarily evaluative task (analogy finding), consistent with prior work showing frontopolar involvement in both evaluation and generation (Bunge et al. 2005; Green, Fugelsang, and Dunbar 2006; Wendelken et al. 2008; Green, Kraemer et al. 2012; Green 2016). Nonetheless, frontopolar cortex is proximate to generally more ventral regions that have been identified with the default mode network, and frontopolar function may well serve to help integrate the products of a putative default mode idea generator and/or integrate default mode network contributions with the category- and rule-based constraints of proximate lateral prefrontal executive regions (Bunge et al. 2003; Cromer et al. 2010).
Within any model of creativity in the brain, it is certain that frontopolar cortex represents only one component/node within a considerably larger network. The present findings are useful in demonstrating gains that can be achieved by stimulating the frontopolar component of creativity-related brain function in the tasks we employed, but future work will be necessary to assess the respective contributions (and potential enhancement) of other candidate nodes identified in emerging models. Future research should also seek to test whether individual differences in trait intelligence and creative abilities might moderate the effects of neuromodulation. The present study's relatively small and homogeneous cohort of high-achieving undergraduates at a selective university limits our ability to adequately address such questions.
Early evidence suggests that tDCS interventions can transiently improve creative performance (Cerruti and Schlaug 2009; Chi and Snyder 2011, 2012; Chrysikou et al. 2013; Simis et al. 2014; Brunye et al. 2015; Colombo et al. 2015; Goel et al. 2015; Mayseless and Shamay-Tsoory 2015), though far more work will be required to distinguish effects on specific cognitive components that support creativity. To our knowledge, the interaction of tDCS by conscious state creativity augmentation has not yet been explored in any creative process. Cerruti and Schlaug (2009) found that anodal, but not cathodal, tDCS applied to left, but not right, dorsolateral prefrontal cortex (DLPFC) elicited better performance on the Remote Associates Test (Mednick 1968), which is sometimes used as a measure of verbal creativity though that study did not involve any cognitive manipulation of state creativity. Colombo et al. (2015) also reported an effect of tDCS on DLPFC, but collapsed analyses across tDCS foci in left and right hemispheres, making anatomical effects difficult to interpret. That study incorporated an implicit cue intended to either constrain or broaden creative thinking in a uses-of-objects task, but the priming manipulation did not have the intended effect (it did not yield a main effect on creative responding). Prime-by-stimulation interactions were not examined, nor were effects of explicit, conscious augmentation of state creativity since the priming manipulation was implicit. Only one study to our knowledge has used tDCS to target frontopolar cortex (Brunye et al. 2015). That study did not employ a cognitive state creativity manipulation and, consistent with our finding of no tDCS effect in the absence of the cue, did not observe an effect of tDCS on semantic distance of verb generation.
In addition to the changes in activity within frontopolar cortex, our recent neuroimaging investigation of augmented state creativity (Green et al. 2015) implicated increased frontopolar connectivity with anterior cingulate cortex (ACC) and right anterior prefrontal cortex activity, as well as increased activity in inferior frontal gyrus (IFG), which is strongly connected with frontopolar cortex. The involvement of these regions is likely to reflect greater demands on response selection/inhibition when forming more creative semantic connections (Barch et al. 2000; Braver et al. 2001; Badre et al. 2005; Hampshire et al. 2010; Gonen-Yaacovi et al. 2013; Aron et al. 2014). Forming a more creative connection requires consideration of a greater number of possible connections and often requires the inhibition of the first response that comes to mind to select a more creative response. Given that tDCS affects not only the area of cortex directly exposed to the electrical field but also the network of distant brain areas connected to the stimulated area (Stagg et al. 2013, Polania et al. 2011), it is probable that the focal frontopolar stimulation administered here had downstream effects on inhibitory centers of the frontal lobe. Such effects are likely to amplify increases in functional connectivity that occur in response to the creativity cue (Green et al. 2015). Indeed, the particularly large effect of tDCS for high-constraint noun prompts in the present study indicates that inhibiting the effect of prepotent associations is a component of the mechanism by which neuromodulation facilitates creativity. The integrative function of frontopolar cortex appears to support the actual formation of creative connections (Ramnani and Owen 2004; Green, Fugelsang, Kraemer et al. 2006; Green et al. 2010; Green and Kraemer et al. 2012; Gonen-Yaacovi et al. 2013), but it is likely that this process occurs in concert with contributions of ACC and other regions that inhibit prepotent responding to make “outside the box” connections more available.
Conclusions
Departing from traditional research treatments of creativity as a static trait, evidence of neural mechanisms that support dynamism in creative performance suggests the encouraging potential of targeted cognitive and neural interventions. Here, we leveraged insights from neuroimaging to target stimulation of a frontopolar region where we predicted that potentiating neuronal activity would facilitate augmentation of state creativity. Results provide novel evidence that tDCS enhances the conscious augmentation of state creativity elicited by cognitive intervention, and extend the known boundaries of tDCS enhancement to analogical reasoning, a form of creative intelligence that is a powerful engine for innovation.
Supplementary Material
Supplementary material can be found at: http://www.cercor.oxfordjournals.org/.
Funding
This work was supported by awards from the National Science Foundation (DRL-1420481), The John Templeton Foundation (#51971), and Pymetrics to A.G.; and the NIH/NCATS via the Georgetown Howard Universities Center for Clinical and Translational Science (KL2 TR000102) to P.T. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Notes
We thank Chris Thomas and Dr Abhishek Datta of Soterix Medical, Inc. for providing electrical field modeling support and renderings for visualization. Conflict of Interest: None declared.
Appendix A
Analogy finding task matrices.


References