





Abstract
The cumulative effects of global change, including climate change, increased population, and more intense industrialization and agribusiness, will likely continue and intensify the course of eutrophication in estuarine and coastal waters. As a result, the symptoms of eutrophication, such as noxious and harmful algal blooms, reduced water quality, loss of habitat and natural resources, and severity of hypoxia (oxygen depletion) and its extent in estuaries and coastal waters will increase. Global climate changes will likely result in higher water temperatures, stronger stratification, and increased inflows of freshwater and nutrients to coastal waters in many areas of the globe. Both past experience and model forecasts suggest that these changes will result in enhanced primary production, higher phytoplankton and macroalgal standing stocks, and more frequent or severe hypoxia. The negative consequences of increased nutrient loading and stratification may be partly, but only temporarily, compensated by stronger or more frequent tropical storm activity in low and mid-latitudes. In anticipation of the negative effects of global change, nutrient loadings to coastal waters need to be reduced now, so that further water quality degradation is prevented.Rabalais, N. N., Turner, R. E., Díaz, R. J., and Justić, D. 2009. Global change and eutrophication of coastal waters. – ICES Journal of Marine Science, 66: 1528–1537.
Introduction
The world's climate has changed, and human activities will continue to contribute to the acceleration of emission of greenhouse gases and rising temperatures. The effects of global climate change (GCC) are already detectable in the decline in snow cover, glaciers and polar ice, which have led to a poleward shift in plant and animal distributions, and caused changes in algal and fish communities and foodwebs (IPCC, 2007). The regional outcomes of the various GCC scenarios will likely manifest in many different and synergistic effects on various components of ecosystems. These outcomes may be difficult to detect in the near term, because they are currently masked by the inherent “background” variability of natural systems, and because there are effects resulting from other human-caused changes (Sanderson et al., 2002; Halpern et al., 2008). The sum of the natural variability, GCC, and human effects is what we refer to as “global change”. The drivers causing coastal eutrophication are set within a larger framework of multiple human-induced stressors, including overfishing, chemical contaminants, coastal habitat degradation, and invasive species (Paine et al., 1998; Breitburg et al., 1999; Rabalais, 2004). For example, the declining salinity and oxygen concentrations prevalent in the spawning areas of the eastern Baltic Sea are not driven by changes resulting only from GCC. The effects of changes in hydrographic conditions, however, are causally related to the reproductive success of cod (Gadus morhua), and when combined with high fishing pressure, such changes resulted in a decline of the stock to the lowest level on record in the early 1990s (Köster et al., 2005). The synergistic effects of GCC and other global effects will make restoration of fish stocks more difficult than currently.
This paper addresses some important mechanisms by which global change may affect the eutrophication of coastal and estuarine ecosystems. Eutrophication, defined as the increased rate of primary production and accumulation of organic matter (modified from Nixon, 1995), usually results from the excessive addition of nutrients, and results in undesirable changes in ecosystems. Eutrophication is a global phenomenon, with significant effects on foodwebs, water quality, and aquatic chemistry (Cloern, 2001; Rabalais, 2004). As in the sense of multiple stressors described above, we are unable to differentiate completely the GCC aspect and the anthropogenic activities aspect of global change on eutrophication. We proceed in the following sequence, however, keeping in mind that there is considerable overlap in the pressures and consequences. We first discuss GCC in the context of physical, chemical, and biological environments, and then with expected landscape changes and uses resulting from increased human pressures. A case study is presented for the northern Gulf of Mexico, which is strongly influenced by the Mississippi River, a river that has already undergone major changes because of human activities and which is likely to be aggravated by GCC. This area, although not unique, is one of the more notable cases of human-caused eutrophication in the global coastal zone.
GCC and coastal systems
If the average global temperature rises by 0.4°C over the next three decades and by 1.8–4.0°C over the next century (IPCC, 2007), there will be major changes in ecosystem structure and function, trophic interactions, and habitat ranges and migration patterns of many species. Most GCC scenarios project predominantly negative consequences for biodiversity and ecosystem goods and services (Figure 1). The major drivers of these changes are increased temperature, sea level rise, enhanced hydrological cycles, and shifts in wind patterns, which might alter coastal currents.
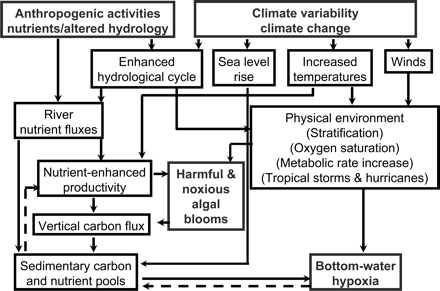
Potential physical and hydrological changes resulting from climate change and their interaction with current and future human activities. The dashed lines represent negative feedback to the system.
Temperature and salinity
Increasing temperatures on their own have the potential to strengthen pycnoclines (density differences dictated by a combination of temperature and salinity) in estuarine and coastal waters, but lower surface salinity (e.g. from increased freshwater run-off) would be more of a factor in stratifying the water column (Figure 2). For example, at 20°C and 32 psu, a 1-psu decline in salinity changes seawater density by a factor of about three relative to a 1°C increase in temperature (Fonfonoff and Millard, 1983). Stronger pycnoclines may result in less diffusion of oxygen from the upper part of the water column to the lower part of the water column, resulting in less dissolved oxygen in bottom waters. Decreasing oxygen concentrations in the lower part of the water column will negatively affect biological communities (Figure 3) and secondary production, and disrupt biogeochemical cycles (Díaz and Rosenberg, 1995, 2008; Rabalais et al., 2001a, 2001b).
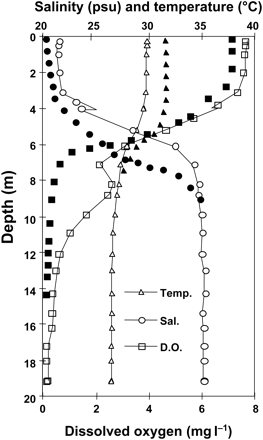
Typical thermocline, halocline, and oxycline for a station in 20 m, ∼100 km west of the Mississippi River delta in midsummer (see Figure 5). The open symbols are recorded data, and the filled symbols are predicted changes with increased surface water temperature and increased freshwater discharge. For dissolved oxygen (D.O.), the predicted response is for lower oxygen in the upper water column because of decreased saturation and lower oxygen higher in the water column and lower values below the thermo- and halocline.
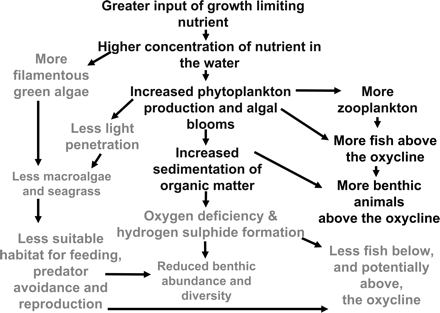
Schematic representation of the cascading effects of increasing nutrients in a coastal ecosystem. The harmful effects of nutrient overenrichment are presented in grey letters.
Regional wind patterns
Increased temperatures may also affect regional wind patterns, resulting in changes in circulation and mixing. If wind patterns or intensity change, coastal currents and their effects on coastal waters might change to either aggravate a low oxygen condition or, conversely, to alleviate it. An example of the former happened off the Oregon and Washington coasts, where wind-driven shifts in the California Current System in 2001, and subsequent years, altered the long-term pattern of upwelling and resulted in extensive hypoxia along the inner shelf (Grantham et al., 2004; Chan et al., 2008).
Tropical storms and hurricanes
If the frequency of tropical storms and hurricanes increases as a result of increased water temperatures in the lower to mid-latitudes as predicted by certain GCC models, the vulnerability of coastal habitats will increase greatly. Warmer Atlantic Ocean sea surface temperatures could increase the frequency and severity of tropical storms (Trenberth, 2005; Hoyos et al., 2006; Kerr, 2006). Recent analyses (reviewed by Kerr, 2008), however, suggested only a modest increase, or even a decrease, in the frequency and intensity of future Atlantic cyclones. Knutson et al. (2007) predicted a modest 18% decrease in the frequency of Atlantic hurricanes by the end of the 21st century and a few per cent increase in the intensity of storms. Conversely, Emanuel et al. (2008) forecasted a few per cent increase in storm frequency and a 7.5% increase in storm intensity.
The effects of hurricanes and tropical storms on stratification, and how much they temporarily alleviate hypoxia in the coastal waters of the northern Gulf of Mexico, depend on several factors (Rabalais et al., 2007a), which include barometric pressure, windspeed, storm trajectory, and the forward speed and circumference of the wind and wave field. The timing of storms is also critical. If hurricanes occur early in the “hypoxia season”, there will only be transient dissipation of hypoxia. If they occur later in summer, restratification may be slowed by an increasing frequency of autumn/winter storms (mixing and thermal turnover). Hurricanes have also caused the development of extensive hypoxia in North Carolina coastal waters and in some Florida systems. The primary factors in lowered dissolved oxygen concentrations after storms in Pamlico Sound, NC (Paerl et al., 2000; Bales, 2003), and Charlotte Harbor, FL (Tomasko et al., 2006), were the higher freshwater loading that resulted in increased stratification and the resuspension of sediments rich in organic carbon from eutrophication in preceding years. Similar situations were observed in the lower Atchafalaya River basin of Louisiana following Hurricane Andrew in 1992, Hurricane Rita in 2005, and Hurricanes Gustav and Ike in 2008.
Biological processes
The rates of biological processes, including photosynthesis and respiration, are expected to increase with higher water temperatures, but only up to a point. For instance, primary production may proceed at a higher rate, but will eventually become limited by light (self-shading) or lack of nutrients. In addition, aerobic organisms will be brought closer to their thermal limits. These two factors, when combined, may result in a decrease in the physiological capacity of aerobic organisms (Pörtner and Knust, 2007).
Enhanced hydrological cycle
Increased precipitation will result in more water, sediments, and nutrients reaching the coastal zone, where they are likely to enhance eutrophication through nutrient-enhanced production, increased stratification, or both (Cloern, 2001; Rabalais, 2004). Reduced precipitation will lower the amount of nutrients and water reaching the coastal zone and, perhaps, result in oligotrophication and reduced fisheries productivity. An example of human-caused reduction in freshwater discharge is the Nile River delta, where water retained by the Aswan High Dam led to a 90% reduction in floodwaters and nutrient loading of the Mediterranean Sea (Nixon, 2003). Fish and shrimp harvests declined by ∼80% after the dam had been constructed. Even in areas with sufficient or increased precipitation, water withdrawal will continue to support human populations and production of food and fibre. The Huanghe's (Yellow River) water discharge to the sea steadily decreased since the 1950s (Yang et al., 1998). From 1990 to 1999, for example, the average water discharge from the Huanghe decreased to 12.3 × 109 m3 year−1, which was only 29% of its discharge in the 1950s (Wang et al., 2005). This was both the result of El Niño/Southern Oscillation events and large dam and reservoir construction on the Huanghe. A large proportion of the world's population is currently experiencing water stress (Vörösmarty et al., 2000), and water diversions are expected to increase.
Sea level rise
The GCC models predict that coastal zones will be exposed to an accelerated sea level rise and increased erosion. Most climate models predict that the eustatic sea level rise during the 21st century will be between 20 and 60 cm (IPCC, 2007). Human actions in the coastal zone have already affected the rate of relative sea level rise by levee and channel construction and withdrawal of water, oil and gas (Morton et al., 2002). Coastal wetlands, including saltmarshes and mangroves, are projected to be negatively affected by sea level rise, especially where they are constrained on their landward side by geomorphology, that is to say, a gradient of increasing elevation or sediment composition, or from the increasing human-induced pressures on coastal areas for space, resources, or pollutant loading, including higher nutrient fluxes. The world's coastal-zone population could grow from 1.2 billion people (in 1990) to between 1.8 and 5.2 billion people by the 2080s, adding to the negative effects of climate change. Coastal wetlands are important ecosystems because of their natural removal of reactive nitrogen (Galloway and Cowling, 2002) and many other pollutants. Decreases in wetland area will result in disruptions in the overall effectiveness of ecosystem services, particularly the removal of excess nutrients that could cause eutrophication. In addition, any degradation of marsh ecosystems associated with the subsequent release of organic matter and nutrients could for a limited time contribute to local instances of eutrophication (Figure 3).
Eutrophication and its symptoms
The causes of eutrophication should not be confused with the process itself. Eutrophication may result from changes in the physical characteristics of a system (e.g. hydrology), biological interactions (e.g. reduced grazing), or an increased organic and inorganic nutrient loading. Although the series of causes may include direct natural or anthropogenic carbon enrichment, eutrophication in the coastal ocean in the 20th and 21st centuries has more often been related to the excessive loading of nutrients, which stimulates phytoplankton growth. Humans are producing more and more reactive nitrogen and phosphorus, resulting in fluxes of nitrogen and phosphorus to coastal waters that far exceed their natural production rate (Bennett et al., 2001; Tilman et al., 2001; Galloway and Cowling, 2002; Foley et al., 2005). Eutrophication is likely to be aggravated where river discharge and fertilizer use increase (Figure 1).
When nutrient-limiting phytoplankton growth is added to an aquatic system, the response of the phytoplankton community is to increase its rate of production and biomass accumulation up to a point (Figure 3). The quantity of fish harvested from an area is positively proportional to the increase in nutrients—but only up to a threshold, after which fish harvests may decline (Caddy, 1993; Nixon and Buckley, 2002; Breitburg et al., 2009). The beneficial aspects of increased primary production are illustrated in black letters in Figure 3, including the increase in fished species and their prey organisms. Negative effects can occur if the increase in nutrients is above the capacity of the system to absorb the increased phytoplankton production. Algal blooms may result in increased sedimentation of organic matter on the seabed and subsequent oxygen depletion when they decompose, especially below the pycnocline (Glibert et al., 2005). Once the oxygen concentration decreases to a critical level, mobile species flee the area, while sessile organisms initiate survival behaviours, or begin to die as dissolved oxygen continues to decline (Rabalais et al., 2001a, b; Díaz and Rosenberg, 2008).
Climate change and increased anthropogenic nutrient loading will make coastal ecosystems more susceptible to the development of hypoxia through enhanced stratification, decreased oxygen solubility, increased metabolism and remineralization rates, and increased production of organic matter. All these factors related to global change may progressively result in an onset of hypoxia earlier in the season and possibly an extended duration of hypoxia, as predicted for Chesapeake Bay (Boesch et al., 2007).
In some shallow-water, well-mixed eutrophic estuaries, the natural diel cycle of dissolved oxygen varies from supersaturation during the day to hypoxia or near-anoxia during the night. The long-term trends (1986–2004) in the well-mixed Skidaway estuary, and in other lower reaches of rivers and estuaries of Georgia, indicate a reduction in both surface and bottom dissolved oxygen saturation (no obvious increase in surface water temperature) that is attributed to increases in ambient concentrations of increased inorganic and organic nutrients, chlorophyll a, bacterial and heterotrophic community metabolism. Calm weather conditions and extended periods of cloud cover (i.e. less light), which reduces production of oxygen by primary producers, often exacerbate the problem of hypoxia in these systems (Verity et al., 2006; Tyler and Targett, 2007).
The prevalence of hypoxia in estuarine and coastal areas is increasing, which is consistent with the idea that hypoxia is largely driven by increased nutrient loading to coastal waters (Díaz and Rosenberg, 2008). The changes in various indices of eutrophication-related hypoxia in the United States demonstrate an increased percentage of estuarine and coastal ecosystems from the 1980s into the 2000s (Table 1). There has been almost a doubling in reports of systems with hypoxia over this 20-year period. It seems quite clear that more and more coastal systems, especially in areas of increased industrialization and mechanized farming, where physical conditions are appropriate and where nutrient loads are predicted to increase, are likely to become eutrophic (with accompanying hypoxia) in the future, even in the absence of GCC.
Percentage of US estuaries with reports of low dissolved oxygen or hypoxia along with the percentage of area and period of the year affected [modified from Rabalais (1998) with the addition of data from Díaz and Rosenberg (2008)].
| 1980s | 1990s | 2000s | ||||
|---|---|---|---|---|---|---|
| Region | Percentage of estuariesa | Percentage of estuariesb | Percentage of areab | Maximum area (km2)b | Monthsc | Percentage of estuariesd |
| North Atlantic | 6 | 22 | 0–2 | 100 | 7–9 | 38 |
| Mid Atlantic | 50 | 59 | 9–22 | 4 500 | 6–9 | 62 |
| South Atlantic | 16 | 62 | 4–12 | 1 150 | 5–9 | 84 |
| Gulf of Mexicoe | 66 | 84 | 12–27 | 8 300 | 6–10 | 91 |
| Gulf of Mexicof | 86 | 32–66 | 20 200 | 4–10 | ||
| Pacific | 21 | 26 | 1–2 | 130 | 8–10 | 55 |
| Total US | 37 | 52e | 8–19e | 14 200e | 5–10 | 64 |
| 53f | 21–43f | 37 400f | ||||
| 1980s | 1990s | 2000s | ||||
|---|---|---|---|---|---|---|
| Region | Percentage of estuariesa | Percentage of estuariesb | Percentage of areab | Maximum area (km2)b | Monthsc | Percentage of estuariesd |
| North Atlantic | 6 | 22 | 0–2 | 100 | 7–9 | 38 |
| Mid Atlantic | 50 | 59 | 9–22 | 4 500 | 6–9 | 62 |
| South Atlantic | 16 | 62 | 4–12 | 1 150 | 5–9 | 84 |
| Gulf of Mexicoe | 66 | 84 | 12–27 | 8 300 | 6–10 | 91 |
| Gulf of Mexicof | 86 | 32–66 | 20 200 | 4–10 | ||
| Pacific | 21 | 26 | 1–2 | 130 | 8–10 | 55 |
| Total US | 37 | 52e | 8–19e | 14 200e | 5–10 | 64 |
| 53f | 21–43f | 37 400f | ||||
aBased on Whitledge (1985).
bBased on Bricker et al. (1999).
cMonths are referred to by number with 1 = January, 2 = February, etc.
dBased on Díaz and Rosenberg (2008).
eDoes not include the Mississippi/Atchafalaya river plume.
fIncludes the Mississippi/Atchafalaya river plume.
Percentage of US estuaries with reports of low dissolved oxygen or hypoxia along with the percentage of area and period of the year affected [modified from Rabalais (1998) with the addition of data from Díaz and Rosenberg (2008)].
| 1980s | 1990s | 2000s | ||||
|---|---|---|---|---|---|---|
| Region | Percentage of estuariesa | Percentage of estuariesb | Percentage of areab | Maximum area (km2)b | Monthsc | Percentage of estuariesd |
| North Atlantic | 6 | 22 | 0–2 | 100 | 7–9 | 38 |
| Mid Atlantic | 50 | 59 | 9–22 | 4 500 | 6–9 | 62 |
| South Atlantic | 16 | 62 | 4–12 | 1 150 | 5–9 | 84 |
| Gulf of Mexicoe | 66 | 84 | 12–27 | 8 300 | 6–10 | 91 |
| Gulf of Mexicof | 86 | 32–66 | 20 200 | 4–10 | ||
| Pacific | 21 | 26 | 1–2 | 130 | 8–10 | 55 |
| Total US | 37 | 52e | 8–19e | 14 200e | 5–10 | 64 |
| 53f | 21–43f | 37 400f | ||||
| 1980s | 1990s | 2000s | ||||
|---|---|---|---|---|---|---|
| Region | Percentage of estuariesa | Percentage of estuariesb | Percentage of areab | Maximum area (km2)b | Monthsc | Percentage of estuariesd |
| North Atlantic | 6 | 22 | 0–2 | 100 | 7–9 | 38 |
| Mid Atlantic | 50 | 59 | 9–22 | 4 500 | 6–9 | 62 |
| South Atlantic | 16 | 62 | 4–12 | 1 150 | 5–9 | 84 |
| Gulf of Mexicoe | 66 | 84 | 12–27 | 8 300 | 6–10 | 91 |
| Gulf of Mexicof | 86 | 32–66 | 20 200 | 4–10 | ||
| Pacific | 21 | 26 | 1–2 | 130 | 8–10 | 55 |
| Total US | 37 | 52e | 8–19e | 14 200e | 5–10 | 64 |
| 53f | 21–43f | 37 400f | ||||
aBased on Whitledge (1985).
bBased on Bricker et al. (1999).
cMonths are referred to by number with 1 = January, 2 = February, etc.
dBased on Díaz and Rosenberg (2008).
eDoes not include the Mississippi/Atchafalaya river plume.
fIncludes the Mississippi/Atchafalaya river plume.
Case history: Mississippi River watershed and Gulf of Mexico
The coupled Mississippi River watershed and coastal Gulf of Mexico ecosystem is a well-known example of continental-scale landscape change, with increased nutrient loads to the gulf resulting from human activity in the watershed (particularly from agricultural practices) and an eutrophic coastal ecosystem manifested by increased primary production and development of extensive and severe seasonal hypoxia (Turner and Rabalais, 1994, 2003, 2004; Rabalais et al., 1996, 2007a, b; Turner et al., 2007). The hypoxic zone in the Gulf of Mexico is the second largest human-caused hypoxic area in global coastal waters (Rabalais et al., 2002). Changes in the watershed, and subsequent flux of nutrients to the gulf, enhance the high phytoplankton productivity that results in noxious and harmful algal blooms and high organic loading to the seabed. The high freshwater discharge supports salinity stratification throughout the year, which is strengthened by warmer surface waters from spring to autumn.
Changes in the Mississippi River watershed
The watershed encompasses >3 225 000 km2 of the North American continent and 41% of the area of the conterminous 48 states of the United States (Figure 4). The river ranks among the world's top ten rivers with regard to length, area of watershed, discharge, and sediment load (Milliman and Meade, 1983). Approximately 58% of the river basin is devoted to agricultural land (Goolsby et al., 1999). In general, average flows peak (22 500 m3 s−1) in March, April, and May, whereas low to average flows (as little as 7000 m3 s−1) occur in late summer and early autumn (Bratkovich et al., 1994). The timing, distribution, and temporal change of discharge volume into the northern Gulf of Mexico affect both the physical oceanography and biological processes leading to seasonal hypoxia.
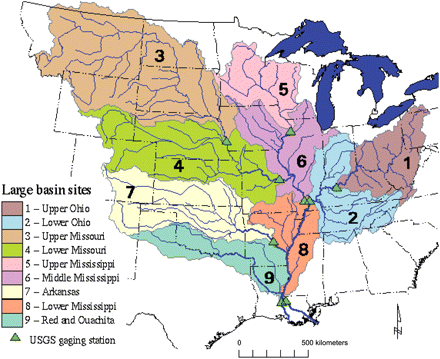
Major sub-basins of the Mississippi River watershed: Upper (basins 5 and 6), Ohio (basins 1 and 2), Missouri (basins 3 and 4), Lower (basin 8), Arkansas (basin 7), and Red (basin 8). Source: Goolsby et al. (1999).
The strongest correlate in all models that predict the size of the northern Gulf of Mexico hypoxic area is the spring nitrate (N) load of the Mississippi River (Scavia et al., 2003; Turner et al., 2005, 2006). Turner et al. (2008) further narrowed the uncertainty in this prediction by adding a year term to account for “legacy carbon” in the seabed and nutrient retention in the watershed. The nitrate load of the Mississippi River (the product of nitrate concentration × discharge) increased ∼300% from the 1950s to the mid-1990s (Figure 4; Goolsby et al., 1999; Goolsby and Battaglin, 2001; Turner et al., 2007). The most significant driver in the change of nitrate load is the increase in concentration of nitrate in the river, not the freshwater discharge (Donner et al., 2002; Justić et al., 2002). Conversely, there is a direct correlation between increases in stream flow and more intense farming practices (Raymond et al., 2008). Orthophosphate loads have not changed, but there has been slight increase in total phosphorus (Turner et al., 2007).
The upper part of the Mississippi River watershed (basins 1–6 of Figure 4) falls within the area of North America predicted to experience increased run-off (higher precipitation) and earlier peak discharge (increased temperature; IPCC, 2007; Milliman et al., 2008). An increase in precipitation (10–30% per century) should result in increased erosion and loss of phosphorus and increased flux of dissolved inorganic nitrogen, particularly nitrates, through the soils and artificially drained agriculture areas. An increase in the base flow of water, because of enhanced seasonal snowmelt, is predicted to result in a 1–2-week earlier peak stream flow. The spring peak delivery of nutrients and freshwater to the northern Gulf of Mexico begins a seasonal cycle of increased primary production and flux of organic matter to bottom waters, which subsequently results in low oxygen concentrations (Rabalais et al., 2007a). The combination of increased nutrient loads (from human activities) and increased freshwater discharge (from GCC) will aggravate the already high loads of nutrients from the Mississippi River to the northern Gulf of Mexico and strengthen stratification (all other factors remaining the same). Higher temperatures in other parts of the watershed are predicted to decrease precipitation and magnify drought conditions. The lower Mississippi River basin is within the sector of North America that should experience lower rainfall in summer, but not severe drought conditions as predicted for the southwestern United States or areas of Africa (IPCC, 2007).
Changes in the coastal Gulf of Mexico
Recent trends for the northern Gulf of Mexico with regard to temperature change indicate that the northeastern quadrant fell within the −0.2 to 0.2°C temperature change for 1970–2004 and that the remaining quadrants fell within the 0.2–1.0°C temperature increase for the same period (IPCC, 2007). These trends are similar to what is predicted for most of the world's oceans, except for the eastern temperate Atlantic, which has a more substantial projected increase of 1.0–2.0°C. The projected average temperature increase in the ocean over the next century is 1.5–2.0°C (IPCC, 2007), which will most likely strengthen the pycnocline in the coastal waters of the Gulf of Mexico (Figure 2). A stronger pycnocline will lower the flux of dissolved oxygen from the upper water column to the lower water column.
Justić et al. (2002, 2003a) examined several climate change scenarios using projected coastal surface water temperatures and flux of nutrients from the Mississippi River watershed (Justić et al., 2003b). They used a coupled physical–biological model representative of a station 100 km west of the Mississippi River, in an area of high-frequency occurrence of midsummer hypoxia, in 20-m water depth and with an average pycnocline at 10 m (Figures 2 and 5; Justić et al., 1996, 1997). The nominal model was based on an observed time-series (1955–2000) for temperature, river freshwater discharge, and nitrate flux. The results of model simulations identified the mid-1970s as a start of the recurring hypoxia in the lower water column (Justić et al., 2002). These results agree well with first reports documenting hypoxia in the northern Gulf of Mexico (Rabalais et al., 2002) and retrospective analyses of sedimentary records for surface water productivity and bottom-water oxygen conditions (Rabalais et al., 2007b). Model simulations that reflected predicted changes in Mississippi River discharge, nutrient concentration, and increasing water temperature were used to calculate the percentage change in the number of years with hypoxia compared with the nominal model. According to the nominal model, there were 19 years that were hypoxic in summer between 1955 and 2000 (Table 2). The number of years with hypoxia for a scenario with pre-1967 nitrate–N concentrations was zero. Miller and Russell (1992) suggested that the average annual discharge of the Mississippi River would increase by 20%, if the concentration of atmospheric CO2 doubled. Similar estimates by Milliman et al. (2008) predict a 20% increase in Mississippi discharge. Scenarios for a 4°C increase in surface water temperature and a 20% increase in freshwater discharge (Miller and Russell, 1992) resulted in an increased frequency of hypoxia of 37 and 32%, respectively. If the increase in temperature and freshwater discharge were coupled, it resulted in a predicted 63% increase in the number of years with hypoxia. Conversely, a 30% decrease in river discharge over the same period (Wolock and McCabe, 1999) resulted in a predicted 58% decrease in the number of hypoxic years. As a simulation of nutrient management strategies that would reduce the concentration of nitrate–N in the Mississippi River, Justić et al. (2003a) applied a 30% reduction in nitrate–N concentration to the nominal model and predicted that the number of years with hypoxia would be reduced by 37%.
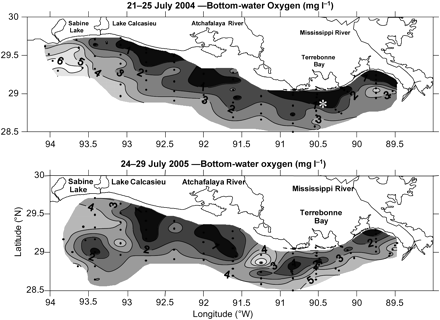
Distribution of bottom-water hypoxia on the Louisiana continental shelf in midsummer 2004 and 2005. The asterisk is the location of station C6C, which is in 20-m water depth, 100 km west of the Mississippi River delta in an area of frequent midsummer bottom-water hypoxia. Source: NNR, LUMCON.
Simulated changes in the frequency of bottom-water hypoxia in the northern Gulf of Mexico in response to various climatic and nitrogen loading scenarios (from Justić et al., 2003b).
| Model scenario | Number of years with hypoxia | Percentage change relative to <2 mg O2 l−1, the nominal model |
|---|---|---|
| Nominal model | 19 | – |
| 1955–1967 N–NO3 | 0 | ∞ |
| −30% Q | 8 | −58 |
| +20% Q | 26 | +37 |
| +4°C | 25 | +32 |
| +20% Q and +4°C | 31 | +63 |
| Management of −30% N–NO3 | 12 | −37 |
| Model scenario | Number of years with hypoxia | Percentage change relative to <2 mg O2 l−1, the nominal model |
|---|---|---|
| Nominal model | 19 | – |
| 1955–1967 N–NO3 | 0 | ∞ |
| −30% Q | 8 | −58 |
| +20% Q | 26 | +37 |
| +4°C | 25 | +32 |
| +20% Q and +4°C | 31 | +63 |
| Management of −30% N–NO3 | 12 | −37 |
The nominal model was forced using time-series of observed monthly values of the Mississippi River discharge (Q) and nitrate–N concentration (N–NO3) over a base period from 1955 to 2000. The scenarios were based on projections of general circulation models for the continental United States, as well as proposed nutrient management plans.
Simulated changes in the frequency of bottom-water hypoxia in the northern Gulf of Mexico in response to various climatic and nitrogen loading scenarios (from Justić et al., 2003b).
| Model scenario | Number of years with hypoxia | Percentage change relative to <2 mg O2 l−1, the nominal model |
|---|---|---|
| Nominal model | 19 | – |
| 1955–1967 N–NO3 | 0 | ∞ |
| −30% Q | 8 | −58 |
| +20% Q | 26 | +37 |
| +4°C | 25 | +32 |
| +20% Q and +4°C | 31 | +63 |
| Management of −30% N–NO3 | 12 | −37 |
| Model scenario | Number of years with hypoxia | Percentage change relative to <2 mg O2 l−1, the nominal model |
|---|---|---|
| Nominal model | 19 | – |
| 1955–1967 N–NO3 | 0 | ∞ |
| −30% Q | 8 | −58 |
| +20% Q | 26 | +37 |
| +4°C | 25 | +32 |
| +20% Q and +4°C | 31 | +63 |
| Management of −30% N–NO3 | 12 | −37 |
The nominal model was forced using time-series of observed monthly values of the Mississippi River discharge (Q) and nitrate–N concentration (N–NO3) over a base period from 1955 to 2000. The scenarios were based on projections of general circulation models for the continental United States, as well as proposed nutrient management plans.
Hypoxia and hurricanes
The 2005 tropical storm season for the Gulf of Mexico is notorious for the devastating effect of hurricanes Katrina (in August) and Rita (in September) on the Louisiana coast. However, two additional earlier storms—hurricanes Cindy and Dennis—generated sufficiently high wave and windfields to disrupt hypoxia on the Louisiana shelf in July, before the scheduled cruise that maps the extent of midsummer hypoxia took place. The subsequent size of the hypoxic area was smaller (11 840 km2) than predicted by the nitrate–N load in May (16 083 km2) based on the Turner et al. (2006) model (Figure 5). However, hypoxia had re-established across a larger area by August (NNR, unpublished data), when Hurricane Katrina crossed the southeastern Louisiana coast. The variability in the changes of oxygen conditions near the bottom in a 20-m water column is illustrated by the 2003 hurricane season (Figure 6). The passage of several tropical storms and hurricanes in June–August 2003 disrupted stratification and hypoxia, but to varying degrees. The path of Tropical Storm Bill was very close to station C6C, but it passed rapidly north and the wave field was insufficient to re-aerate the bottom waters. Although it passed well to the south of station C6C, Hurricane Claudette generated a field of 30-knot winds at the site of the observing system, as it moved slowly towards the west. As with Hurricanes Cindy and Dennis in 2005, which resulted in a smaller area of hypoxia than predicted for the spring nitrate–N load, the effect of Hurricane Claudette in 2003 was sufficient to reduce the size of the bottom-water hypoxia to 8560 km2, which was ∼2.5% less than the predicted size of 20 000 km2. Tropical Storms Erika and Grace had opposite effects. The former had no effect on hypoxia former, whereas the latter caused an increase in bottom oxygen.
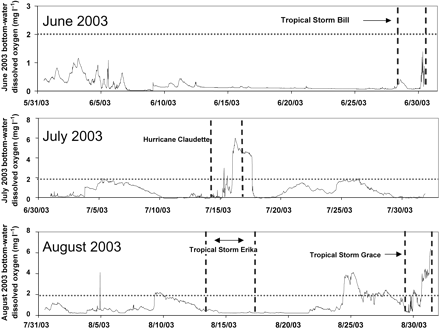
Bottom-water dissolved oxygen concentrations at 20-m water depth at station C6C (Figure 5) for June–August 2003, with the timing of tropical storms and hurricanes superimposed. The horizontal dotted line at 2 mg l−1 represents the operational definition of hypoxia. The vertical dashed lines delimit the temporal extent of various tropical storms and hurricanes. Source: NNR, LUMCON.
Summary
There is little doubt that human population growth and human activities have altered global climate, landscapes, hydrological cycles, and the flux of nutrients essential to plant growth at accelerating rates over the past several centuries (Vitousek et al., 1997; Galloway and Cowling, 2002; Galloway et al., 2003; IPCC, 2007). The continued and accelerated export of nitrogen and phosphorus to the world's coastal ocean, even without climate change, is the trajectory to be expected unless societal intervention in the form of controls or changes in culture are successfully pursued. The largest proportional increases in nutrient loading are predicted for southern and eastern Asia, and are associated with population growth, subsequent increased fertilizer use, and increased industrialization (Seitzinger et al., 2002). These excess nutrients are finding their way to the coastal zones in increasing amounts, especially during the last half of the 20th century. The increased production of biofuels in many countries will further amplify nutrient delivery from the land to the sea (National Research Council, 2007; Runge and Senauer, 2007; Simpson et al., 2007).
Coastal water quality is currently on the decline. The projected increased nutrient loading is predicted to increase the incidence and severity of eutrophication and hypoxic water formation. The likelihood of strengthened stratification alone, from increased surface water temperature as the global climate warms, is sufficient to worsen hypoxia where it currently exists and will facilitate its formation in other coastal areas. The interplay of increased nutrients and stratification may be offset by the potential for more frequent and/or severe tropical storm systems. The future activity of tropical storms under increased global temperatures, however, is unclear, remains the subject of debate, and only temporarily alleviates hypoxia.
The overall forecast is for more eutrophication and for hypoxia to worsen, with increased occurrence, frequency, intensity, and duration. The need remains for water and resource managers to reduce nutrient loads to reduce global eutrophication and its associated negative effects and to prevent further degradation.
Acknowledgements
We thank the National Oceanic and Atmospheric Administration, Center for Sponsored Coastal Ocean Research, NGOMEX06 grant numbers NA06NPS4780197 to the Louisiana Universities Marine Consortium and Louisiana State University and NA06NOS4780141 to LSU for support. This is publication number 104 of the NOAA CSCOR NGOMEX program.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}