




Abstract
The ability to form anticipatory representations of ongoing actions is crucial for effective interactions in dynamic environments. In sports, elite athletes exhibit greater ability than novices in predicting other players’ actions, mainly based on reading their body kinematics. This superior perceptual ability has been associated with a modulation of visual and motor areas by visual and motor expertise. Here, we investigated the causative role of visual and motor action representations in experts’ ability to predict the outcome of soccer actions. We asked expert soccer players (outfield players and goalkeepers) and novices to predict the direction of the ball after perceiving the initial phases of penalty kicks that contained or not incongruent body kinematics. During the task, we applied repetitive transcranial magnetic stimulation (rTMS) over the superior temporal sulcus (STS) and the dorsal premotor cortex (PMd). Results showed that STS-rTMS disrupted performance in both experts and novices, especially in those with greater visual expertise (i.e. goalkeepers). Conversely, PMd-rTMS impaired performance only in expert players (i.e. outfield players and goalkeepers), who exhibit strong motor expertise into facing domain-specific actions in soccer games. These results provide causative evidence of the complimentary functional role of visual and motor action representations in experts’ action prediction.
INTRODUCTION
Mounting research evidence has shown that action perception is strictly linked to motor representations (Prinz, 1997; Hommel et al., 2001). Indeed, effective interactions in dynamic environments require the prediction of the outcome of perceived actions and the formation of anticipatory representations of motion sequences. This ability has been either attributed to general visual processes, also responsible for the perception and recognition of environments and their elements (such as objects; e.g. Zago and Lacquaniti, 2005), or to a unique system specialized in the perception of the movements of conspecifics (Verfaillie and Daems, 2002; Ramnani and Miall, 2004; Wilson and Knoblich, 2005). Accordingly, several neuroimaging and neurophysiological studies have documented that the ‘action observation network’ (AON) includes not only visual, occipito-temporal areas, but also motor, fronto-parietal areas (Rizzolatti and Craighero, 2004; Fadiga et al., 2005; Grafton, 2009; Van Overwalle and Baetens, 2009). In this view, subjective experience has been considered to serve a crucial role in the recognition and simulation of ongoing actions (Hecht et al., 2001; Casile and Giese, 2006; Aglioti et al., 2008; Urgesi et al., 2012). Yet, the involvement of visual or motor brain representations in experts’ action perception has to be clarified.
Neuroimaging evidence has indicated that motor experience can significantly modulate the extent of the activation of the AON. For example, viewing dance movements activated the AON more in professional dancers than in novices (Calvo-Merino et al., 2005, 2006; Cross et al., 2006, 2009a,b). When controlling, however, for the relative contribution of motor and visual expertise with the observed dance moves, only the fronto-parietal, but not the temporal visual areas, were specifically modulated in motor experts (Calvo-Merino et al., 2006; Cross et al., 2009a,b). Moreover, the responses of neurons in the superior temporal sulcus (STS), a critical node of the AON, are influenced by previous action perception (Jellema and Perrett, 2003), suggesting that these neurons may use visual experience to form a representation of ongoing actions (Perrett et al., 2009). This representation is purely perceptual, as STS neurons do not respond during action execution (Rizzolatti and Craighero, 2004). Conversely, neurons responding to both action observation and execution in the premotor cortex (mirror neurons; di Pellegrino et al., 1992; Gallese et al., 1996) may allow using previous motor experience with similar actions for predicting the future of ongoing actions, and thus building internal anticipatory models of even briefly perceived actions (Wilson and Knoblich, 2005; Gazzola and Keysers, 2009; Avenanti and Urgesi, 2011; Friston et al., 2011).
A significant example of the need for anticipatory representations of ongoing actions is in the case of sports. Athletes in time-demanding sports have to plan their actions based on the future of perceived movements executed by their opponents in the minimum amount of time; thus, an accurate prediction of the outcome of observed actions is deemed as necessary for successful performance. Indeed, previous research with elite athletes has shown that they own a unique ability to predict the future of opponents’ actions. For example, in various sports it has been found that both expert athletes and observers are able to provide earlier and more accurate predictions of the outcome of sport actions, compared to novices; however, while expert observers, such as coaches, base their predictions on the initial ball trajectory, elite athletes rely more on the perceived body kinematics of their opponents (Abernethy et al., 2008; Aglioti et al., 2008; Tomeo et al., 2012; Urgesi et al., 2012). Furthermore, these athletes’ superior perceptual abilities are associated with differential activations in the motor cortex (Aglioti et al., 2008; Tomeo et al., 2012) and in body-related visual areas (Abreu et al., 2012) during observation of domain-specific actions. While these findings provide indications about the involvement of both visual and motor representations in the experts’ superior abilities for predicting the fate of observed actions, no study has so far provided causative evidence about their relative functional roles.
To dissociate the role of visual and motor action representations in experts’ action perception, in the present study we investigated whether the suppression of visual and motor areas in experts had different detrimental effects with respect to novices. We applied a temporal occlusion paradigm (adapted from Tomeo et al., 2012), in which the presentation of soccer penalty kicks was interrupted at the football contact, thus only providing information on the kicker’s body kinematics. The kicks could be directed to the left or to the right side of the goalpost, but in half of the trials the videos were manipulated so that an incongruent football contact followed the initial body running phase. At the offset of each video, we asked outfield players, goalkeepers and novices to predict the actual outcome of the kick (i.e. ball placed to the left or to the right of the goalpost). In keeping with the results of Tomeo et al. (2012), presenting the whole body movement up to the football contact ensured that all groups, independently of their expertise, had enough information to make correct predictions of the kick outcomes. Thus, all groups were expected to have comparable performance at baseline in this paradigm, while expertise-related improvements of prediction performance are stronger when only the initial body kinematics cues are available (Aglioti et al., 2008; Tomeo et al., 2012; Urgesi et al., 2012). Nevertheless, we expected that according to relative motor or visual expertise the three groups may use different perceptual cues and types of processing, more linked to motor simulation or to visual processing of body kinematics. Such different processing patterns should require different involvement of motor and visual areas that are activated during observation of body actions. Thus, to test the relative causative role of motor and visual action representation in experts and novices, during observation of the video clips, we applied active or sham repetitive transcranial magnetic stimulation (rTMS) over two critical nodes of the AON; namely the left dorsal premotor cortex (PMd) and left STS. Left hemisphere areas were targeted because we presented right foot penalty kicks, which are represented in the left motor cortex, and because the left hemisphere AON seem to be dominant in action observation independently from the side of the observed movement (Caspers et al., 2010). We hypothesized that STS-rTMS would impair task-performance in all groups, with greater detrimental effects for those with more visual expertise (i.e. goalkeepers). Conversely, PMd-rTMS should impair performance of soccer players but not novices, with greater effects for those players who exhibit greater motor expertise (i.e. outfield players).
MATERIALS AND METHODS
Participants
The experiment sample consisted of 16 expert soccer outfield players aged 18–37 years (mean = 23.56 years, s.d. = 5.24), 16 expert goalkeepers aged 18–39 years (mean = 23.13 years, s.d. = 5.44) and 16 novices aged 19–28 years (mean = 21.77 years, s.d. = 1.96). No differences in age were detected between the participants’ groups (one-way ANOVA, [F(2,45) = 0.72, P = 0.49, ηP2 = 0.03]. All the participants were men and right-handed according to a standard handedness inventory (Briggs and Nebes, 1975). Four outfield players, one goalkeeper and three novices reported left-foot dominance, while the remaining 40 participants reported right-foot dominance. Outfield players trained for a mean of 6.88 h/week (s.d. = 1.36) and had played soccer for a mean of 16.44 years (s.d. = 6.27). Goalkeepers received training for a mean of 7.75 h/week (s.d. = 2.14) and had played soccer for a mean of 15.13 years (s.d. = 4.97). The two expert groups did not differ for either h/week (t(16) = −1.34, P = 0.191) or years of practice (t(16) = 0.636, P = 0.53). All outfield players and goalkeepers played in amateur Italian Soccer League teams, while all novices reported no experience of having received training or playing soccer in teams. All participants reported normal or corrected-to-normal visual acuity in both eyes and were naïve to the purpose of the study. Informed consent was obtained from all participants, and they were compensated with 25€ for taking part. The experimental procedures were approved by the ethics committee of the Scientific Institute ‘E. Medea’ and complied with the ethical standards of the Declaration of Helsinki (1964). Before taking part in the experiment, all participants had to complete a medical questionnaire, screening for neurological and other medical problems, as well as other contraindications to TMS (as described in Wassermann, 1998; Rossi et al., 2009).
Stimuli and apparatus
Stimuli were adapted by those used in Tomeo et al. (2012) research study. These were all video clips derived from digitally recorded videos of a male expert soccer outfield player (playing in an Italian Amateur Soccer League team) performing a series of penalty kicks under the instructions to place the ball at about 2.5–3.5 m to the left or to the right side of the goal center. The videos were recorded from the frontal plane, with the camera placed at a height of 150 cm and at a distance of 11 m from the goal line, corresponding to the actual position of the goalkeeper. Eight videos were used: four displaying left-directed initial running and left kicks and four displaying right-directed initial running and right kicks (with reference to the observer’s perspective). Each video lasted 800 ms and it was split in 20 frames by using Adobe Premiere software (Adobe Systems Incorporated, San Jose, CA), with the last frame being the football contact point. Each video was presented in both congruent and incongruent conditions. In congruent condition, each video clip was presented with its own original total number of frames, thus displaying the initial running phase and a congruent football contact scene in the last frame. In incongruent condition, the initial running phase was combined with the last frame displaying an incongruent football contact scene (i.e. a kick in the opposite direction with respect to that indicated by the initial running phase). By this manipulation we managed to obtain eight congruent (four right- and four left-directed) and eight incongruent action video clips (four with left-directed initial running and right-directed football contact; four with right-directed initial running and left-directed football contact; see Figure 1 for example of stimulus manipulation).
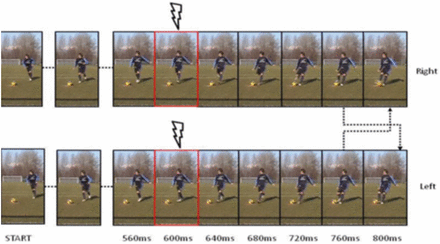
Sequence of video clips presentation in a typical trial. The arrows denote the change of the last video frame for the control of congruent and incongruent trials. The flash symbol and highlighted frame indicate the start of delivering the TMS pulses.
During the experiment, participants seated in a dimly light room in front of a 19-inches CRT monitor (resolution of 1027 × 768 pixels, refresh frequency at 75 Hz), in which videos were presented on a black background and subtended a 14.4° × 11.5° region of optical view. The presentation of the stimuli and the TMS triggering were controlled by E-Prime version 2.0 software (Psychology Software Tools Inc., Pittsburgh, PA). The same software also controlled for the randomization of the stimuli within experimental blocks and the recording of participants’ responses.
Transcranial magnetic stimulation
Online rTMS was performed by means of a 70 mm figure-of-eight stimulation coil (Magstim Double 70-mm Air Film Coil), connected to a Magstim SuperRapid2 Stimulator (The Magstim Company, Carmarthenshire, Wales), producing a magnetic field up to 0.8 T at the coil surface. The location of the primary motor cortex ‘hot spot’ for activating muscles of the right hand was determined prior to the main experiment. This was achieved by trial and error exploration relative to its typical location, with single-pulse TMS applied at a low rate (<0.2 Hz). Surface Ag/AgCl cup electrodes (1 cm diameter) were placed in a belly-tendon montage and connected to a Magstim MEP Pod module (sampling rate, 6000 Hz; band-pass filters, 20 Hz–2 kHz). First, we identified the scalp location that consistently gave rise to motor-evoked potentials (MEPs) with the highest amplitude in the first dorsal interosseous (FDI) hand muscle. Then, we determined the lowest stimulation intensity that, during rest, evoked MEPs with amplitude higher than 50 μV on at least 50% of occasions (the ‘resting motor threshold’).
The online rTMS protocol applied in this study involved five pulses at 10 Hz at 120% of the participant’s resting motor threshold. This online, high-frequency protocol has been identified to disrupt the functions related to the target area (Walsh and Pascual-Leone, 2003). The stimulation sites (Figure 2) on each participant’s scalp were identified by means of a SofTaxic Navigator system (EMS, Bologna Italy). Skull landmarks (nasion, inion and two preauricular points) and ∼60 points providing a uniform representation of the scalp were digitized by means of a Polaris Vicra Optical Tracking System (NDI, Canada). Coordinates in Talairach space (Talairach and Tournoux, 1988) were automatically estimated by the SofTaxic system based on an MRI-constructed stereotaxic template. The dorsal premotor area (PMd) was targeted in the left precentral gyrus (x = −50, y = −1, z = 44; Cross et al., 2011) corresponding to Brodmann’s area 6, whereas the STS area was targeted in its posterior aspect (x = −62.9, y = −52.5, z = 9.4; Caspers et al., 2010) corresponding to Brodmann’s area 21. The coil position during the experiment was monitored by means of the SofTaxic Navigator system, which also estimated the projections of the stimulation sites on the brain surface. The coil was held tangentially to the scalp, with the handle pointing at 45° backwards/laterally for PMd stimulation and 45° backwards/upwards for STS stimulation. Sham stimulation was delivered by tilting the TMS coil 90° over the PMd area; this procedure allows to fake stimulation with the same noise and scalp contact as during real stimulation but no current reaches the brain (Rossi et al., 2009). Sham stimulation was applied over the scalp location of only PMd and not of STS to reduce the number of experimental conditions, as we did not expect any effect at all of sham stimulation on performance wherever it was applied.
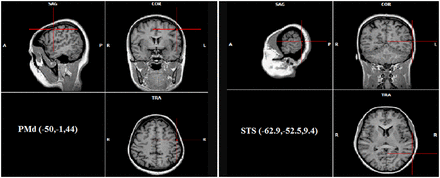
Stimulation sites applied in the current study.
Procedure and design
Each experimental session lasted ∼90 min and consisted of three blocks, with one block for each stimulation site. In each block, 96 trials were presented (6 repetitions × 16 video clips) in a fully randomized order. The order of blocks was counterbalanced between subjects. A small break was allowed between blocks (Figure 3). A trial started with the presentation of a centrally located fixation cross for 1 s, which was followed by the experimental videos presented for 800 ms at the center of the computer monitor. The rTMS pulses were delivered 600 ms after the onset of each video clip (see also Figure 1). Thus, the rTMS train lasted from 200 ms before the video offset to 200 ms afterwards. At the end of each video presentation, a prompt frame appeared asking the participants to press with their right index or middle finger the left or right button of the computer mouse to indicate whether the kick displayed in the video clip was directed to the left or right side, respectively. Participants were instructed to be as fast as possible in their responses while maintaining accuracy. Presenting the prompt frame before allowing participants to respond avoided response anticipations and ensured that all participants in all conditions viewed the whole video clip before responding. This, however, prevented us from considering latencies as measure of processing efficiency in perceiving the stimuli. Thus, our main dependent variable was response accuracy (see also Tomeo et al., 2012). At the end of each experimental session, all subjects were debriefed; none reported any problems and/or major discomfort due to TMS.
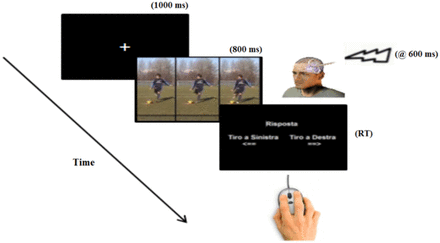
Sequence of a typical trial in the study.
Kinematic analysis of the visual stimuli
To identify the kinematic differences between the left- and right-directed initial running phases and kicks, we calculated the angles formed by the model’s upper and lower limb joints at two frames: 200 ms before and at the football contact point. The definition of the joint angle profile was performed using dedicated software for motion analysis (Dartfish Connect v. 4.0, Dartfish Ltd., Fribourg, Switzerland). For each frame, we defined the amplitude of the joint angles for the right hip, left hip and right side of the waist (Figure 4). For each joint, the mean angle values of the four left and the four right kicks were compared in each of the two frames by series of non-parametric independent tests (Mann–Whitney U Test).

Example of body kinematic measurements. Grey lines denote the joint angles, for which we took measurements.
Behavioral data analysis
The percentage of correct responses (accuracy) was calculated for each participant in each experimental condition. For the incongruent video clips, the correctness of responses was defined on the basis of the direction indicated by the last available cue (football contact point). Furthermore, as a supplementary analysis, to directly compare the magnitude of the interferential effects in the three groups, a percent index of rTMS effect when compared with Sham was computed for each stimulation site with the following formula: (Accuracy active rTMS–Accuracy Sham)/(Accuracy active rTMS+Accuracy Sham). The use of such a difference-by-sum ratio procedure (which is analogous to a quotient computation) allowed us to scale the estimation of the rTMS effects to the individual baseline performance. Finally, as in two-alternative-forced-choice tasks, like the one in the present study, it is possible that accuracy percentage conflates bias with decision, we also calculated and analyzed d′ and response bias scores (natural logarithm of β; ln β), to validate and further expand our findings. For statistical analyses we used repeated-measures and mixed-model analysis of variance (ANOVA) models implemented in Statistica 8 software (StatSoft, Inc, Tulsa, OK), after checking that normality and sphericity assumptions were not violated. All post hoc pairwise comparisons were performed using the Duncan test. A significant threshold of P < 0.05 was set for all analyses. Effect sizes were estimated using the partial eta square measure (ηP2).
RESULTS
Action kinematics
The most critical kinematics difference (Figure 5) in the transition from the running to the football contact phase of both left and right action videos was identified between the angles formed by the right side of the model’s waist. More specifically, for congruent videos (left running to left football contact, right running to right football contact), a significant increase of the joint angle was observed for both sides (left: U = 1, Z = −2.02, P < 0.05; right: U = 0, Z = −2.31, P < 0.05). This increase was also significant for the incongruent transition from right running to left football contact (U = 0, Z = −2.31, P < 0.05), whereas it was not observed for the incongruent trials showing left running and right football contact (U = 4, Z = −1.15, P = 0.25). Thus, with respect to waist angle changes, the incongruent trials depicting right running and left football contact appeared similar to congruent trials, whereas this was not the case for the incongruent trials depicting left running and right football contact. Furthermore, for the right hip angles, we found a significant decrease for both congruent (left: U = 0, Z = 2.31, P < 0.05; right: U = 0, Z = 2.31, P < 0.05) and incongruent videos (left: U = 0, Z = 2.31, P < 0.05; right: U = 1, Z = 2.02, P < 0.05). Finally, for the left hip, no significant differences were identified for both congruent and incongruent videos.
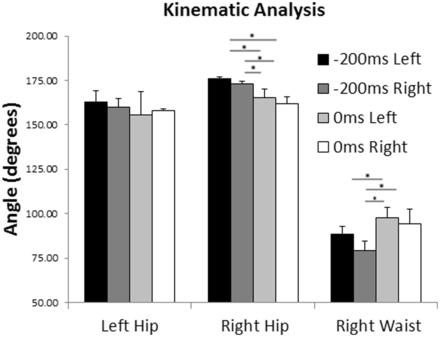
Joint angles data. Error bars denote standard errors.
Baseline behavioral performance
On the basis of the aforementioned kinematic analysis results and to check whether the sides of initial running and kick (left, right) affected participants’ responses, we first run a mixed-model ANOVA on the accuracy data (untransformed) for the Sham condition, with group (outfield players, goalkeepers and novices) as a between-subjects factor, and type of action (congruent, incongruent) and side of initial running (right, left) as within-subjects effect. The ANOVA results revealed only a significant two-way interaction [F(1,45) = 26.33, P < 0.001, ηP2 = 0.37] showing that left-side initial running/right kicks were responded better than right-side initial running/left kicks in the incongruent condition (P < 0.001). The main effect of group and the two- and three-way interactions involving the group factor were not significant [all F(2,45) < 1.36, P > 0.26, ηP2 < 0.57], showing comparable group performance at baseline. We thus tested how TMS over STS and PMd affected the performance of the three groups while all of them could perform the task at baseline.
rTMS effects
Accuracy
Considering the significant differences between left and right sides of initial running and kicks, we run separate ANOVAs for the accuracy values (untransformed) of each initial running side condition, with group (outfield players, goalkeepers, novices) as a between-subjects factor, and type of action (congruent, incongruent) and stimulation (STS, PMd, Sham) as within-subjects effects (Figure 6). For the right-side initial running condition, we found only a significant main effect of action type [F(1,45) = 750.41, P < 0.001, ηP2 = 0.94], with better performance for congruent than incongruent trials. No other main effects or interactions were significant [all F < 1]. For the left-side initial running condition, the ANOVA revealed significant main effects of stimulation [F(2,90) = 17.66, P < 0.001, ηP2 = 0.28] and action type [F(1,45) = 480.46, P < 0.001, ηP2 = 0.91]. Also, the two-way interactions between group and action type [F(2,45) = 4.57, P < 0.05, ηP2 = 0.17] and between group and stimulation [F(2,90) = 18.2, P < 0.001, ηP2 = 0.29] were significant, and were further qualified by a significant three-way interaction between group, type of action and stimulation [F(4,90) = 2.48, P < 0.05, ηP2 = 0.1].
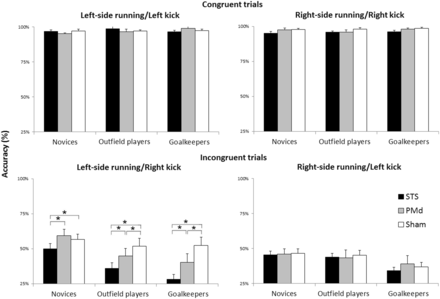
Accuracy data (untransformed) in the task. Error bars denote standard errors.
To explore the significant three-way interaction for left-side initial running, we run separate ANOVAs for each of the three groups, with action type and stimulation as within-subjects effects. For the group of novices, the ANOVA revealed a significant main effect of action type [F(1,15) = 133.74, P < 0.001, ηP2 = 0.9] and a significant two-way interaction [F(2,30) = 3.63, P < 0.05, ηP2 = 0.19]. Post hoc tests revealed that novices’ performance for incongruent trials was significantly impaired in the STS with respect to both PMd (P = 0.005) and Sham (P = 0.029) rTMS conditions, between which in turn it did not differ (P = 0.384). For the group of outfield players, the ANOVA revealed significant main effects of action type [F(1,15) = 143.71, P < 0.001, ηP2 = 0.91] and stimulation [F(2,30) = 7.04, P < 0.005, ηP2 = 0.32]. Also, the two-way interaction between action type and stimulation was significant [F(2,30) = 10.7, P < 0.001, ηP2 = 0.42] showing that outfield players’ performance was significantly impaired in the STS than PMd (P = 0.003) and Sham (P < 0.001) stimulation conditions; importantly, however, also PMd-rTMS had a detrimental effect with respect to Sham (P < 0.001). In a similar vein, the ANOVA for goalkeepers showed significant main effects of action type [F(1,15) = 211.78, P < 0.001, ηP2 = 0.93] and stimulation [F(2,30) = 9.68, P < 0.001, ηP2 = 0.39], as well as significant two-way interaction [F(2,30) = 8.58, P < 0.005, ηP2 = 0.36]. Post hoc tests revealed that STS-rTMS impaired performance with respect to PMd-rTMS (P = 0.005) and Sham (P < 0.001); then again, PMd-rTMS impaired performance as compared to Sham (P = 0.006).
Magnitude of the rTMS effect.
Although the performance of all groups in predicting incongruent left-side initial running/right kicks was interfered by STS-rTMS, the magnitude of the effect (transformed data: Figure 7) for novices was significantly lower with respect to goalkeepers (t(30) = 2.63, P = 0.013), but not to outfield players (t(30) = 1.79, P = 0.083). On the other hand, the magnitude of the effects of PMd-rTMS was significantly lower in novices than both goalkeepers (t(30) = 2.16, P = 0.039) and outfield players (t(30) = 2.14, P = 0.04). No difference was obtained between outfield players and goalkeepers (both P > 0.18).
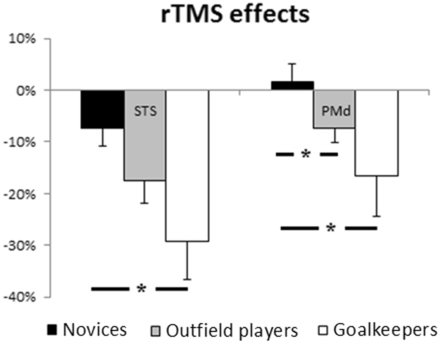
rTMS effects (transformed accuracy data) between experimental groups. Error bars denote standard errors.
Signal detection theory analysis.
As mentioned before, in two-alternative forced choice tasks, like the one we applied at the present study, accuracy data may be affected by systematic response strategies. Because the present study design was aimed at testing how the ability of three different groups of observers to predict the outcome of congruent and incongruent soccer actions was affected by rTMS applied over STS and PMd, performance could be affected not only by sensitivity to detect an incongruence between the initial running phase and the football contact, but also by systematic bias to respond in accordance with the direction indicated by the initial running phase. To control for these effects and validate our accuracy data findings, we used the signal detection theory to calculate d′ and ln β. We did this by plotting our accuracy data, either for left- or right-side initial running videos, according to presence vs absence of incongruence between the initial running phase and the football contact; the proportions of left-right responses for each condition were coded according to whether they corresponded to the side of the initial running phase (i.e. no incongruence was detected) or to the opposite side (i.e. incongruence was detected). The d′ and ln β scores were then calculated, considering the proportion of ‘incongruence-detected’ responses in the incongruent condition as ‘hits’ and the proportion of ‘incongruence-detected’ response in the congruent condition as ‘false alarms’; separate calculations were performed for the videos displaying left and initial running directions (Figure 8). Then, we run a repeated-measures ANOVA on these data with group (outfield players, goalkeepers, novices) as a between-subjects factor, and stimulation (STS, PMd, Sham) and side of initial running (left, right) as within-subjects effects.
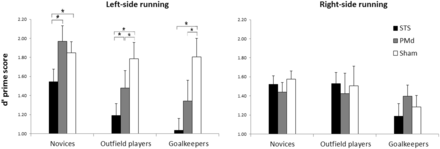
d’ prime scores in the task. Error bars denote standard errors.
For the d′ scores, the ANOVA revealed main effects of stimulation [F(2,90) = 11.44, P < 0.001, ηP2 = 0.2] and initial running side [F(1,45) = 5.92, P < 0.05, ηP2 = 0.12], a two-way interaction between stimulation and side of initial running [F(2,90) = 12.65, P < 0.001, ηP2 = 0.22], further validated by a three-way interaction between group of subjects, stimulation and initial running condition [F(4,90) = 2.75, P < 0.05, ηP2 = 0.11]. To explore the significant three-way interaction, we run separate ANOVAs for each of the three groups, with stimulation and initial running side as within-subjects effects. For the group of novices, the ANOVA revealed a significant main effect of initial running side [F(1,15) = 8.26, P < 0.05, ηP2 = 0.35] and a significant two-way interaction [F(2,30) = 4.44, P < 0.05, ηP2 = 0.23]. Post hoc tests revealed that novices’ performance for left-side initial running was significantly impaired in the STS with respect to both PMd (P = 0.003) and Sham (P = 0.02) rTMS conditions, between which in turn it did not differ (P = 0.356). For the group of outfield players, the ANOVA revealed a significant two-way interaction between stimulation and initial running side [F(2,30) = 7.98, P < 0.01, ηP2 = 0.35] showing that outfield players’ performance for the trials depicting left-side running was significantly impaired in the STS than PMd (P = 0.019) and Sham (P < 0.001) stimulation conditions; importantly, however, also PMd-rTMS had a detrimental effect with respect to Sham (P = 0.015). Finally, the ANOVA for goalkeepers showed a significant main effect of stimulation [F(2,30) = 6.45, P < 0.01, ηP2 = 0.3], as well as a significant two-way interaction [F(2,30) = 6.04, P < 0.01, ηP2 = 0.29]. Post hoc tests revealed that for the left-side initial running condition, STS-rTMS impaired performance with respect to Sham (P < 0.001) and marginally compared to PMd-rTMS (P = 0.06); then again, PMd-rTMS impaired performance as compared to Sham (P = 0.005). No significant differences were observed between stimulation conditions for the trials showing right-side initial running in all three groups.
Finally, for ln β scores the ANOVA revealed non-significant main effects or interactions [all F < 2.38, P > 0.13, ηP2 < 0.065], with overall bias greater than 1, pointing to a moderate tendency to report more often incongruent running and football contact cues in all conditions and groups. Thus, the effects of rTMS on performance were not mediated by change in response bias.
DISCUSSION
In the present study, we investigated how the action prediction performance of expert soccer players (outfield players and goalkeepers) and novices was affected by rTMS interference with visual (STS) and motor (PMd) areas. The results showed that, for congruent actions, all three groups achieved ceiling effects in performance and, thus, no difference was observed between experts and non-experts. However, the presentation of an incongruent football contact significantly impaired the performance of all participants, especially when the videos showed left-directed running and right football contact (kick). Kinematics analysis, indeed, showed that the body kinematics in the transition from right running to left football contact was compatible with a congruent action. This left participants with minimal information for discriminating the actual kick direction, and they tended to base their responses on the initial running phase (performance<50%). Conversely, the transition from left running to right football contact altered the actual kinematics of the kick, and this allowed participants to detect incongruent body-kinematics cues and predict with relatively better accuracy the actual outcome of the kick. Although the loose clothes of our model player, which were similar to soccer team kits used in matches, may have prevented an accurate measurement (and perception) of subtle kinematic cues, the alterations detected by our analyses were large enough to be perceived during video presentation. Importantly, it was in the incongruent condition that rTMS interference with STS and PMd impaired performance as compared with Sham stimulation. In particular, while STS-rTMS impaired action discrimination performance in all three groups, PMd-rTMS had an effect only in those groups with direct motor (and visual) expertise with the displayed, domain-specific actions (i.e. outfield players and goalkeepers).
Previous research studies have shown that motor experts present superior perceptual abilities in reading the body kinematics of observed actions (Farrow and Abernethy, 2003; Aglioti et al., 2008; Urgesi et al., 2012). This allows them to predict earlier and more accurately the outcome of others’ actions. In line with these studies, Tomeo et al. (2012) found that expert outfield players and goalkeepers outperformed novices with regards to predicting the outcome of observed congruent penalty kicks after observation of the initial only running phase. On the other hand, when also the football contact was presented, the results were in line with the present study, indicating a comparable high-level performance in all expert and novice participants. Hence, while sport experts have superior action-prediction abilities with respect to novices, this perceptual advantage is specific for reading initial body kinematics. On the other hand, the availability of crucial visual information extracted by scenes, such as the football contact and the initial ball trajectory, can also give an advantage and increase the performance of novices in predicting the fate of on-going actions. In this view, visual and motor experience may play different, complementary roles in action prediction (Urgesi et al., 2012). Indeed, visual experience may foster visual action representations that are used to describe and to understand the visual dynamics of the movements and of the related contexts. In contrast, motor experience may allow for motor, simulative, body-kinematics-based representations that are used to predict and to anticipate the future actions of other individuals (Wilson and Knoblich, 2005; Abernethy and Zawi, 2007; Schütz-Bosbach and Prinz, 2007; Smeeton and Huys, 2010; Urgesi et al., 2010).
The main aim of the present study was to test the effects of interfering with the visual and motor nodes of the AON in experts and novices. Previous studies (Calvo-Merino et al., 2005, 2006; Cross et al., 2006, 2009a, b) have shown that the activity of these two nodes are differently affected by visual and motor expertise and that this modulation is associated with experts’ greater ability in understanding others’ actions (Aglioti et al., 2008; Calvo-Merino et al., 2010). Beyond this correlational finding, which cannot rule out that the association between experts’ higher motor activation and superior perceptual abilities is just epiphenomenal (Avenanti and Urgesi, 2011; Avenanti et al., 2013a, b), the present study provides causative evidence that the functional role of PMd in action perception is dependent on direct motor experience with the observed actions. Indeed, while for both experts and novices a significant impairment of performance was observed following interference with STS, interference with PMd activity impaired only outfield players’ and goalkeepers’ performance.
It has been suggested that the activity of PMd during action observation reflects the inner simulation of the ongoing actions, enabling the observer to create anticipatory representations of perceived known actions (Grezes and Decety, 2001; Avenanti et al., 2007; Urgesi et al., 2007, 2010; Stadler et al., 2011). In line with this notion, we can estimate that suppression of the PMd area in our expert players impaired their performance in the task compared with Sham stimulation, as they were deprived from the ability to depend on their motor experience to create internal anticipations for the outcome of the perceived actions. In the present study, this was evident only for the case of incongruent actions and not for the congruent ones, thus being specific for perceptual tasks in which the observers had to dynamically update the internal action representations on the basis of new and contrasting perceptual evidence. After all, it is in the case of incongruent, ambiguous or incomplete actions that continuous motor information serves a crucial role into accurately perceiving the relevant cues and completing the missing information using internal (motor) models of the spatial and temporal deployment of the actions. However, as mentioned before, all subjects achieved ceiling effects in predicting congruent penalty kicks, and thus any differences between the stimulation conditions may not be obvious.
Recent neurophysiological and neuroimaging research has indicated that the AON may be organized as an active feed-forward system. In particular, STS nodes are thought to pass computations and information from the visual areas to the network (Nishitani et al., 2004); accordingly, stimulation of left (van Kemenade et al., 2012) and right (Grossman et al., 2005) STS interferes with action processing, and patients with lesion of STS present disorders in biological motion perception (Saygin, 2007). Thus, STS seems critical for action perception. Importantly, however, suppression of the STS nodes results in a compensatory increase of action simulation (Wilson and Knoblich, 2005; Avevanti et al., 2013a, b), suggesting that more motor simulation is required when perceptual action processing is more blurred. In the present results, we observed that predicting incongruent actions was significantly decreased following STS inhibition as compared with both Sham and PMd stimulations. Thus, after suppression of STS all participants, and especially goalkeepers, tended to respond more on the basis of the initial kinematics of the running phase and did not detect the incongruent football contact. According to the aforementioned theory, that could be due to the fact that by suppressing STS a greater weight was given to anticipatory action models based on the inner motor simulation of the initial body kinematics. However, that proved detrimental for the present task as simulation of the initial kinematics deprived the subjects from the ability to recognize the following incongruent cues and, thus, adjust their responses. Importantly, such detrimental effect was greater for goalkeepers than novices probably reflecting their greater visual experience with frontal views of penalty kicks as compared with both novices and outfield players. On the other hand, no difference was obtained between the effects of PMd TMS for goalkeepers and outfield players on their ability to predict front-facing actions, in keeping with studies showing that observation of actions viewed from both third-person (front) and first-person (back) perspective engenders activation of fronto-parietal areas (e.g. Alaerts et al., 2009; Vingerhoets et al., 2012; Wiggett et al., 2012). Accordingly, previous behavioral studies (Sebanz and Shiffrar, 2009; Cañal Bruland et al., 2010) have shown better detection of fake actions viewed from the front than side way in both players and goalkeepers as compared with individuals with no specific experience with the actions, although the front viewing perspective is more customary for goalkeepers than for players.
The present results may shed light on the neural bases of the ability to detect deceptive behaviors in others. Indeed, although we did not ask our model to intentionally deceive the observers, our manipulation of the congruence between the running phase and the football contact introduced body-kinematics incongruence, which characterizes deceptive actions (Kunde et al., 2011). Indeed, effective deception implies: (i) providing exaggerated body-related cues that induce others to make incorrect action predictions, and (ii) minimizing or delaying postural cues that may inform others of possible sudden changes (Brault et al., 2010). Thus, the situation is highly reminiscent of successfully fake moves, in which expert kickers can alter the kick direction up to 174 ms before the football contact (van der Kamp, 2006). Detecting deceptive action, thus, requires identifying the incongruence between honest and bluffing body-kinematics cues and flexibly updating ongoing action representation on the basis of upcoming information, abilities which seem to require the motor nodes of the AON (Tidoni et al., 2013).
Previous research has reported mixed results with regards to the role of experience into detecting deceptive actions. For example, studies with elite athletes have shown that motor experts are better into detecting deception compared with less experts or novices (i.e. Jackson et al., 2006; Cañal-Bruland et al., 2010), whereas in other cases it has been reported that the athletes’ superior ability to read the opponents’ body kinematics can disadvantage them into detecting when they are trying to deceive them (Ripoll et al., 1995; Dessing and Craig, 2010). In the Tomeo et al. (2012) study, it was found that outfield players, with respect to goalkeepers and novices, were more susceptible to be fooled by the incongruence between the initial body kinematics and the initial ball trajectory. When coupled with the earlier prediction abilities of outfield players and goalkeepers, with respect to novices, this finding suggests that outfield players automatically create anticipatory representations of perceived actions based on the observation of initial body kinematics and, thus, they are more prone to be fooled. Of course, the same applies to expert goalkeepers, with the difference that they can inhibit these internal anticipatory representations and thus, update them when incongruent perceptual cues are present. Accordingly, using single-pulse TMS to measure cortico-spinal excitability, Tomeo et al. (2012) showed that motor facilitation during observation of incongruent vs. congruent actions was lower for goalkeepers, higher for novices, and comparable for outfield players, indicating that predicting the actual outcomes of fooling actions requires updating simulative motor representations based on visual representation. Although in the present study we did not find any difference between the three groups’ baseline performance in detecting the incongruence between the running phase and the football contact, such performance was differently affected by rTMS interference with the PMd area, which was more detrimental for outfield players and goalkeepers than for novices. Conversely, although interference with STS impaired the performance of all groups, goalkeepers were mostly affected, suggesting their greater relying on visual, and not only, motor action representations (Tomeo et al., 2012).
Overall, in the present study we tried to investigate the causative role of visual and motor action representations on the experts’ ability to predict the actual outcome of ongoing actions on the basis of incomplete and incongruent perceptual information. Though these trials were manipulated in a way that made kick presentation to look less naturalistic, the results clearly indicated that both experts and non-experts under specific circumstances can predict the actual outcome of familiar or unfamiliar actions and that the inclusion of incongruent body kinematics dramatically affects their performance. Most importantly, though, experts and non-experts seem to use different neural mechanisms in this task, as it was illustrated by the different effects of suppressing the visual and motor nodes of the AON in this study. Although both experts and novices can access to visual action representations in STS, only experts are equipped and use internal motor representations to predicts others’ behavior. Further studies in the field could include a repertoire of every-day actions, in which people exhibit different levels of expertise, as well as actions depicting intentional deception and/or some level of incoherence. This way, the present findings could be further validated to confirm and disentangle the relative contribution of visual and motor experience in the formation of action representations in visual and motor areas.
The research was supported by grants from Istituto Italiano di Tecnologia SEED 2009 (Prot. no. 21538; to C.U.), from the Ministero Istruzione Università e Ricerca (Progetti di Ricerca di Interesse Nazionale, PRIN 2009; Prot. no. 2009A8FR3Z; Futuro In Ricerca, FIR 2012, Prot. N. RBFR12F0BD; to C.U.], and from Istituto di Ricovero e Cura a Carattere Scientifico ‘E. Medea’ (Ricerca Corrente 2013, Ministero Italiano della Salute; to C.U.). The authors thank Tommaso Zentilin for his help in expert soccer players’ recruitment and data collection.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}